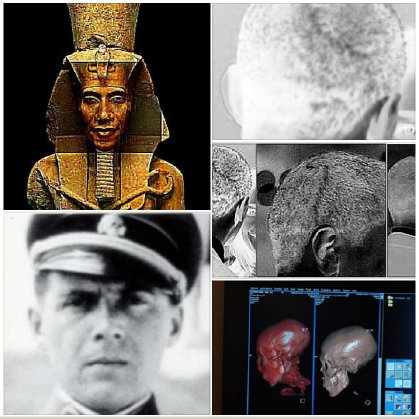


O'Akhenaten, O-Bama & Mengele: First published in 2012. A case for the "CLONING" the "son of perdition".
A "BASTARD CHILD" of HELL IS "BORN"... Born of a whore. THE (mAN of Lawlessness) may be the hideous creation of "ARTIFICIAL INSEMINATION" from the Ancient DNA of Akhenaten. THIS SON of PERDITION, "whose father IS the devil, carries out his father's desires. And like his father, who was a murderer from the beginning, not holding to the Truth, for there is no Truth in him. When he lies, he speaks his native language, for he is a liar like his father, the father of lies".
Hussein's fraudulent birth certificate was designed to not just cover "WHERE" he was born, but to ALSO not disclose "WHO" his genetic paternal, "father" honestly is. The "COVER-UP" will give credence to the future claim by Francis, the false prophet of Rome, that this "anti-Christ" is the "PROOF", (evidence) of the superior race of the extra-terrestrial genetic "fatherhood" an alien origin of human race.
The "man of lawlessness" is the appointed representative of an "extra-terrestrial" entity claim of human "origin" who has returned to earth to "save their own creation" from demise. As "man" will be presented to humanity, whose "paternal", (of the male genetic origin) is undocumented (not known) and whose mother, (maternal) genetic origin), is of a "whore" by nature, the exact opposite of Yeshua Messiah! This appointment by Hell will manifest at the time of the coming global government "alien among us disclosure". MOST of humanity will embrace the idea, thus Satan's GREAT DECEPTION, that will rob Heaven of scores of souls, who will follow this BEAST straight to Hell.
Terra firma (of earth) vs Extraterrestrial (Originating from outside of the Earth's atmosphere, from space, or from another planet; alien to Earth or its environment). Believers will need to cling to every Word of YHWH to overcome the lies that are manifesting about the human origin. Genesis specifically proclaims the "man" is the exclusive creation of YHWH taken from "TERRA FIRMA", (EARTH). Remember this when the proclamation is that man is from genetic alien extraterrestrial origin!!!
"the LORD God formed man out of the clay of the ground (terra firma) and blew into his nostrils the breath of life, and so man became a living being.
O'Akhenaten, O-Bama & Mengele:
*** Between April and May 1960, Dr. Mengele was confirmed to have been in Buenos Aires. Just before or after the capture of SS Eichmann, Mengele disappeared off the radar map. Obama II was born in August 1961. Normally, it would mean that he was conceived in December 1960 during the period of Dr. Mengeles most secretive disappearance episode during the early 1960s. In fact, his trail remained cold until he reportedly surfaced in Brazil in the late 1960s.
Born August 4, 1961, Barry Soetoro, who changed his name to Barack Hussein Obama continues to Keep his Past Identity cloaked in a Shroud of Secrecy. Dr. Ben Carson says he wants President Obama to come clean about the shroud of secrecy surrounding his past, including his birth records.
On November 24, 1982, while Obama II was a student attending Columbia University, Barack Obama, Sr. Was found dead in Kenya. The cover story is that Obama, Sr. Drove his car into a tree following a day of drinking in Nairobi. However, Obama, Sr.,s family, sister, Hawa Auma Obama, and his friend Patrick Ngei believe that he had been assassinated, Obama didnt die out of pure accident . Bright Luos were eliminated by the Kenyatta government.
So, what did Stanley Ann really mean when she suggested to her friends that she could bear children without dating and marriage? Her comment suggests an unusual acute awareness for 1960 of the existence of in vitro fertilization, embryo transfer, and artificial insemination technology- positive eugenics.
Out of everyone in the world, SS Dr. Josef Mengele and the underground SS Knights of the Black Sun had been the most secretive, treacherous and satanic forces that would have conceived such a diabolical plan.
Dr. Mengele disappeared during the time of conception and birth of Obama II. During that time, Dr. Mengele was secretly deeply involved in clandestine artificial insemination in human beings. When Obama II and Stanley Ann returned to Hawaii in 1962, Dr. Mengele was exposed by Argentine historian Jorge Camarasa artificially creating the master race of blond haired, blue eyed Aryans in twins by the dozens in Candido Godoi, Paraguay. Barry Soetoro, (Hussein Obama) was just part of the experimental process of the CIA and was raised with intent and purpose always within their grasp and oversite.
Dont forget pursuant to the SS SCIENTIFIC SATANIC PRINCIPLE of the DIVINE POWER of OPPOSITES that if they had the power to create the MASTER RACE in duality, they had to power to create the SUBSERVIENT.
They had the money, technology, resources, clandestine support, international networks and motive to put a covert Diabolical Black Human Cyborg in the Whitehouse to continue the 1000 Year THIRD REICH and its Satanic Principle of Aryan Planet Domination and the Annihilation of People of Color. An African-American Brother? You never seen anything like him. KEEP DREAMING THAT THE BOYS FROM BRAZIL IS A WORK OF PURE SCIENCE FICTION- AND THAT FAT MEAT AIN'T GREASY!
Dr. Josef Mengele and the SS developed Artificial Insemination and Fertilization that even still today THE CULT OF THE SS stand at the pinnacle and forefront of Modern Genetic and Reproductive Technology. (VN: and that alone should make you damn nervous about the future existence of the human race, if they do away with all of us and only have their clones to deal with, then they are safe and exposed and can live like animals.
Beyond Lebensborn, Himmlers Secret - CIA- All in the Family:
British Intelligence Advisor: CIA Conducted DNA Test on Obama Found No Match to Alleged Grandparents.
Stanley Anns father, Stanley Armour Dunham, had been Obama, Sr.s CIA handler. Stanley Anns mother, Madelyn Dunham, above, also managed the agencys secrets. In 1960, she started working at Hawaii's largest bank, Bank of Hawaii, and was promoted to be one of the first female bank vice presidents in 1970. Madelyn Dunhams expertise had been in the banks escrow accounts department described to be primarily used for 3rd party money transfer accounts between two people who basically do not trust each other and still need a deal to be done. Madelyn handled the secret escrow-payoff accounts of the CIA front company, Bishop Baldwin Rewald Dillingham Wong (BBRDW), Ltd.
BBRDW was primarily run by Ronald R. Rewald. Rewald was a CIA agent recruited to spy on the student radical movement in America in the 1960's. Now Bishop Baldwin expanded its operations greatly. Satellite offices opened up in more than a dozen cities worldwide. It now employed a staff of nearly 200 people. Rewald lived in a Hawaiian estate near Diamond Head valued at over a million dollars in 1980. Bishop Baldwin had a fleet of cars and a chauffeur to drive around Rewald and Bishop Baldwins clients. The company which had four accounts at its incorporation in 1979 had 110 by 1983. And in such exotic places as Hong Kong and the Cayman Islands.
Rewald was now meeting with people like the Sultan of Brunei and Vice-President George Bush and arranging secret arms deals with Rajiv Gandhi of India. In fact, this last seems to have been the major CIA use of the company I.e. To spirit weapons and arms of all types into Pacific Rim countries. Bishop Baldwin also used businessmen to collect intelligence and to direct the flow of capital into American companies. It also was used as a cover for more sinister assignments like the assassinations of leftist leaders and sympathizers.
BBRDW and Rewalds cover was blown in Hawaii by a tip to a news reporter in 1983. Just prior to their cover being blown, CIA Special Agent Jon S. Lodeesen just happened to show up at their doorstep January 12, 1983 for a special assignment in Hawaii.
During the Falkland Islands War of April 1982 between Argentina and Britain, Ron Rewald met with the Gehlen Orgs SS/CIA representatives in Uruguay, Licio Gelli of the Masonic Lodge P2, Klaus Barbie and Heinrich Rupp, to arrange for the Argentine purchase of the French-made Exocet missile used in war.
BBRDW and Rewald had also been a pipeline for the Reinhard Gehlen Org. Madelyn Dunham retired from the Bank of Hawaii in 1986, so it is highly likely that Stanley Anns mother, Madelyn Dunham, handled the Gehlen Orgs secret transfer account to buy Exocet Missiles through the Bank of Hawaii. Stanley Ann Dunham had been a multi-generational asset deeply embedded in the Reinhard Gehlen SS-CIA fraction.
The perfection of positive eugenics was a top-secret mission of the THIRD REICH and a clandestine project of Reichsfuhrer SS-I Heinrich Himmler and the SS. Even today, two of Himmlers SS doctors stand at the pentacle of Human Fertilization and Embryology to Fabricate Aryans from scientific genetic material; and in duality (Satanic Principle) Annihilate non-Aryan genetic material.
(VN: given what we have since learned, it may well be the other way around. Remember, the gov to this day is taking the DNA of newborn babies and following and tracking RH Neg citizens without any reason given. We covered much here about RH negs and they are the ones that are Aryan, and Hitler was obsessed with them and their blood negative is found nowhere in nature. Hmmm, wonder what that means? A different species?).
Himmlers top in vitro fertilization and artificial insemination specialists were SS Teutonic-Knights of the Black Sun and neither one of them were tried at Nuremberg. One was captured in 1945 by the Soviets and may have been turned into a Soviet mole in West Germany, and the other escaped justice under the shroud of the Gehlen Org and Operation CAMPUS.
In 1961-1962, this agency doctor was in Brazil with Lodeesen and Jim Jones of the Peoples Temple. This particular doctor was known to be involved in clandestine artificial insemination and in vitro fertilization projects. He is the subject of the popular 1978 movie thriller, The Boys From Brazil.
Dr. Mengele and Obama II: Timeline of the BOY FROM BRAZIL.
Jim Jones idolized Dr. Josef Mengele. There is cause to believe that while in Brazil in 1961, Jones had met his mentor and had been initiated into the CULT OF THE SS. Additionally, Jones Peoples Temple mind control experiments were an extension of Mengeles Auschwitz experiments.
At Mendocino State Mental Hospital where the entire staff was dismissed and replaced by Jim Joness personnel, according to mind-control survivors, Dr. Mengele had been there in association with Jim Jones programming patients and inmates. It is reported that Dr. Mengele also visited Jonestown, Guyana.
On May 11, 1960, the Israeli MOSSAD captured Obersturmbannführer SS Otto Adolf Eichmann, one of the major organizers of the Final Solution in San Fernando, an industrial community north of the center of Buenos Aires.
MOSSAD agents had been in Argentina since April 1960. At the time, Dr. Mengele was also discovered to have been living in Buenos Aires, Argentina. MOSSAD agents found Dr. Mengeles address and actually took pictures that they determined beyond any doubt the identity of the notorious Demon of Death of Auschwitz. The MOSSAD walked away from Dr. Mengele, one of the worlds most wanted Nazi war criminals. I gather that he was a little too big for their boots protected by U.S. National Security like SS Baron Otto von Bolschwing.
*** Between April and May 1960, Dr. Mengele was confirmed to have been in Buenos Aires. Just before or after the capture of SS Eichmann, Mengele disappeared off the radar map. Obama II was born in August 1961. Normally, it would mean that he was conceived in December 1960 during the period of Dr. Mengeles most secretive disappearance episode during the early 1960s. In fact, his trail remained cold until he reportedly surfaced in Brazil in the late 1960s. [87]
Now, you should have some idea where Stanley Ann could have got her notions of having children without natural procreation-beyond Lebensborn and why Jim Jones showed up in Hawaii. She had been primed for the job as a multi-national CIA/MK ULTRA asset by the satanic (Negative Eugenics) plans of Dr. Josef Mengele; and Jim Jones showed up in Hawaii to dust the trail of his mentor and master.
Dr. Mengeles Land of Aryan Twins:
In a recent book, Mengele: The Angel of Death in South America, the Argentine historian Jorge Camarasa, a specialist in the post-war Nazi flight to South America, contends that Dr. Mengele re-surfaced in about 1962 or early 1963 in the enclave of Colonias Unidas, Paraguay that was founded by German immigrants in 1900.
From there in 1963, Dr. Mengele began to make regular trips to another predominantly German community just over the border in Brazil the farming community of Candido Godoi. (VN: Oh, how very interesting, that coincides with the fact that Bush Jr, and Rev Moon, just bought up the top half of Paraguay and since these cretins do nothing for just one reason, I am wondering if this fraud was one of the reasons? The Cloning of a US President and the need to bury any evidence of what may well have happened there?).
In Candido Godoi, people remembered that Dr. Mengele talked of artificial insemination in human beings, and that he continued working with animals, proclaiming that he was capable of getting cows to produce male twins.
Nobody knows for sure exactly what date Mengele arrived in Candido Godoi, but the first twins were born in 1963, the year in which we first hear reports of his presence, Camarasa said. I think Candido Godoi may have been Mengeles laboratory, where he finally managed to fulfill his dreams of creating a master race of blond haired, blue eyed Aryans, he said.
Dr. Mengele also worked out of the infamous post-war compound of Cologna Dignidad near Paral, Chile founded in 1961 by Nazis straight out of Germany. The UN commission found that the above Dictator for life General AUGUSTO PINOCHETs military intelligence agency DINA-sponsored torture chambers at Colonia Dignidad.
Its underground bunker and chambers were used as research center to continue Dr. Mengeles work of refining the art of torture, prisoners have allegedly been subjected to different experiments without any interrogation Prisoners charge that torture is personalized through an initial interrogation which establishes the personal traits of the individual This data is then used to program the torture sessions so that the result is a totally debilitated person who will comply with any demand.
According to a UN report, in Colonia Dignidad there is a specially equipped underground torture center with small soundproofed cells, hermetically sealed. The detainees heads are covered with leather hoods, which are stuck to their faces with substances that are supposedly chemicals. In these cells, interrogations are carried out through electronic equipment, including loudspeakers and microphones, while detainees are tied naked to metal frames to receive electric shocks.
(VN: oh, wow, now Gitmo makes a lot more sense than it ever did before and given the results where a man was so mind controlled he agreed to blow up his own balls in his pants, and not say a word, then you know he was mind controlled. I told you these people are absolutely insane, and add in the bloodline issue, they are doubly insane. We have to shut down GITMO FOR THE SAKE OF HUMANITY. We are guilty if we let this continue in our name as the people of this country.)
The CIA Medical Experimental Compound at Jim Jones field station and agricultural commune in Jonestown, Guyana had been established as a U.S. Shadow of Cologna Dignidad. Jeannie Mills, a white overseer and ring leader of the Peoples Temple that had been assassinated in Berkeley in 1980, claimed to have seen films taken inside a Chilean torture camp while at Jonestown. These films were either taken at the Colonia Dignidad or another located at Pisagua. At that time, the only source possible for the films had to have been the CIA, DINA or Dr. Mengele.
Contrary to HollyWeird and government propaganda, Dr. Mengele didnt appear to be a frightened fugitive constantly on the run from the law. To the contrary, after the capture, conviction and execution of Obersturmbannführer SS Otto Adolf Eichmann in 1962, the Great Nazi Satanist appeared to have travelled freely throughout South America, Canada, Germany and the United States without a care in the world polarizing the populations, spreading death, destruction and CHAOS.
In 1985, Senator Alfonse M. DAmato (R-N.Y.) testified at a Senate hearing that the U.S. Government had evidence from Cables and other documents indicate that Mengele had a network available to him in raising large sums of money and that high officials (in South America) gave him protection
In other words, Dr. Mengele didnt die in 1979 and was right in the middle of the CIA Iran-Contra CRACK Cocaine Scandal. Of course, U.S. Government agencies claimed they didnt know what Senator DAmato was talking about, because the CIA had also been protecting and supporting him since it was formed in 1947.
Out of everyone in the world, SS Dr. Josef Mengele and the underground SS Knights of the Black Sun had been the most secretive, treacherous and satanic forces that would have conceived such a diabolical plan.
Dr. Mengele disappeared during the time of conception and birth of Obama II. During that time, Dr. Mengele was secretly deeply involved in clandestine artificial insemination in human beings. When Obama II and Stanley Ann returned to Hawaii in 1962, Dr. Mengele was exposed by Argentine historian Jorge Camarasa artificially creating the master race of blond haired, blue eyed Aryans in twins by the dozens in Candido Godoi, Paraguay.
Dont forget pursuant to the SS SCIENTIFIC SATANIC PRINCIPLE of the DIVINE POWER of OPPOSITES that if they had the power to create the MASTER RACE in duality, they had to power to create the SUBSERVIENT.
They had the money, technology, resources, clandestine support, international networks and motive to put a covert Diabolical Black Human Cyborg in the Whitehouse to continue the 1000 Year THIRD REICH and its Satanic Principle of Aryan Planet Domination and the Annihilation of People of Color. An African-American Brother? You never seen anything like him. KEEP DREAMING THAT THE BOYS FROM BRAZIL IS A WORK OF PURE SCIENCE FICTION- AND THAT FAT MEAT AIN'T GREASY!
In Part II, I intend to show where, when and how Dr. Josef Mengele and the SS developed Artificial Insemination and Fertilization that even still today THE CULT OF THE SS stand at the pinnacle and forefront of Modern Genetic and Reproductive Technology. (VN: and that alone should make you damn nervous about the future existence of the human race, if they do away with all of us and only have their clones to deal with, then they are safe and exposed and can live like animals.
The resulting frogs expressed the donor genetic markers, thus proving that the frogs were derived from donor nuclei and not oocyte nuclei left behind from faulty enucleation.
Many other investigators in the United States, England, France, China, and Japan soon entered the field. They too confirmed the totipotency of blastula nuclei in other frog species as well as in some salamander species (reviewed in Di Berardino 1997a).
The pioneers of nuclear transplantation concluded in a classic understatement that although the method of nuclear transplantation should be valuable principally for the study of nuclear differentiation, it may also have other uses (Briggs and King 1952, p. 462). We discuss this research and other applications of cloning below. In retrospect, it is clear that the amphibian nuclear transplantation procedure became the prototype for cloning multicellular animals.
Although the method of nuclear transplantation should be valuable principally for the study of nuclear differentiation, it may also have other uses. (Briggs and King 1952, p. 462)
Clearly, 1997 was the year of the clone (Figure 1). The cover illustration of Nature (27 February 1997) announced the birth of Dolly, the ewe cloned from an adult sheep in Scotland, and Science (19 December 1997) proclaimed Dolly to be the breakthrough of the year. Newspapers, news magazines, radio, and television were even more fervent in their reports of Dolly.
Even now, several years later, Dolly (Wilmut et al. 1997) and a number of mice (Wakayama et al. 1998, Wakayama and Yanagimachi 1999) and calves (Kato et al. 1998, Wells et al. 1999), all cloned from adult cells, continue to evoke fascination. Was Dolly the first animal to be cloned? Of course not. Why, then, the sudden, almost unprecedented attention to the announcement of a cloned sheep? We believe that because Dolly was the first animal cloned from an adult cell, she stimulated scientists, theologians, ethicists, journalists, and politicians to contemplate the application of cloning to humans.
The point of this article is not to reconsider the extension of cloning to humans, a subject that has already been covered (e.g., Silver 1997, Kolata 1998, Nussbaum and Sunstein 1998), but to consider the genuine rationales that stimulated the original and continuing efforts in cloning research. Cloning was never intended as a procedure for the simple multiplication of animals.
Frogs are cheap in the United States, as are sheep in Scotland. The reasons for cloning are more complex than simply producing identical animals, and in this article, we consider those reasons and the results that have been obtained with the procedure. We place cloning in the historical context of developmental biology and review results obtained with the procedure. Some readers may wonder how two scientists with cloning experience view the ethics of human cloning. We offer our views in the epilogue.
Cloning's roots in nineteenth-century biology:
How do cells become specialized during development? One cell, the zygote, gives rise to a multiplicity of cells that in time become increasingly specialized, or differentiated. In the latter part of the nineteenth century, August Weismann believed that differentiation results from the differential and sequential partitioning of the genome as the cells divide (reviewed by Wilson 1928, Spemann 1938).
The attractiveness of the now-discarded Weismann hypothesis was that it could be tested. Wilhelm Roux, in an 1888 experiment (Spemann 1938), killed one cell (blastomere) of a two-cell amphibian embryo and found that a half-embryo developed, suggesting that some genes are lost during cell replication. However, in 1892, Hans Driesch found that if the blastomeres of two-cell sea urchin embryos were physically separated, entire embryos formed from each blastomere (Spemann 1938); similar results were obtained when amphibian blastomeres were isolated, provided that they contained a portion of cytoplasm known as gray crescent material. Thus, the genome was not diminished, but rather reproduced during cell division. (Roux's half-embryos were later interpreted to result from the inhibitory effect of the dead blastomere.)
CLONING HAS PROVIDED INSIGHTS INTO NUCLEAR DIFFERENTIATION, NUCLEAR REPROGRAMMING, CELLULAR AGING, AND GENOMIC IMPRINTING:
Blastomere separation of embryos beyond the 2- to 16-cell stage (depending on species) was noninformative for testing genomic potential because the cells had too little cytoplasm. But in 1894, Jacques Loeb fortuitously observed that fertilized sea urchin eggs sometimes ruptured when exposed to hypotonic solutions (Spemann 1938).
The extruded portion of the egg was usually bereft of a nucleus and remained uncleaved. Occasionally, nuclei traversed the isthmus between the cleaving egg and the extruded cytoplasmic material. In such cases, the extruded cytoplasm cleaved along with the main mass, resulting in the formation of two entire embryos. In 1914, Hans Spemann performed a conceptually identical experiment on an amphibian egg (Spemann 1938).
He constructed a zygote with a noose made of baby hair, causing the egg to assume the shape of a dumbbell. When the cleaving (nucleated) portion reached the 8- or 16-cell stage, he loosened the constriction and permitted a nucleus to move to the non-nucleated cytoplasmic portion. Here, too, the non-nucleated portion cleaved and formed a clone of its nuclear donor.
These primitive nuclear transplantation (cloning) experiments affirmed the view that the complete genome is replicated during cell division, at least during early cleavage. The stage was now set for modern cloning experiments to examine the genomic potential of older embryonic cells. However, Spemann (1938) could see no way for the moment to manually insert a nucleus from older embryos into enucleated cytoplasm. Robert Briggs and Thomas J. King found a way.
Success in Philadelphia:
Around 1943, Briggs (1911-1983), an embryologist working at what is now the Fox Chase Cancer Center in Philadelphia, wanted to determine whether the genome of older somatic nuclei remains equivalent to the zygote nucleus throughout development (Patterson n.d.). One possible approach was to transplant a somatic nucleus into an oocyte whose own nucleus had been removed and then observe what type of development occurred.
Briggs was aware that Comandon and de Fonbrune (1939) had transplanted single nuclei in the unicellular amoeba, but he recognized that nuclear transfer in the oocytes of metazoans would be more arduous because of their complex physiological and biochemical requirements for embryonic development.
Over the next 7 years, while analyzing the development of haploid and triploid frog embryos, he became experienced with various microsurgical techniques that would contribute to the procedure of nuclear transfer. In 1948, one of us (Di Berardino) joined Briggs's laboratory and was there to witness the first cloning of metazoan animals in 1952.
In 1949, Briggs began searching for a research fellow to develop a nuclear transplantation procedure for the North American leopard frog, Rana pipiens. He also applied to the National Institutes of Health for funds to support the project. His first attempt was rejected because the reviewers considered his proposed research a harebrained scheme with little chance of success
However, a second application was successful, and Briggs brought King in as his research fellow in 1950. The two scientists first determined that eggs lacking a functional nucleus but containing a cell division center developed at best into partial blastulae (Briggs et al. 1951). This baseline study established the total developmental potential of the recipient host; any further development could, therefore, be credited to an introduced nucleus.
Briggs and King's nuclear transfer procedure involved first activating an oocyte at metaphase II of meiosis by pricking it with a glass needle, which initiated the metabolic changes normally induced by the sperm. Approximately 15 minutes later, a pit (black dot) appearing on the surface of the oocyte was micro surgically removed with another glass needle; this surgery removed the oocyte's chromosomes, resulting in an enucleated egg. Finally, a blastula cell was aspirated into a glass micropipette whose lumen was slightly smaller than the diameter of the cell to gently break the cell membrane.
The broken cell was then transferred to the animal hemisphere of the enucleated egg, which permitted the nucleus to interact with the cytoplasmic molecular milieu of the oocyte and to undergo nuclear reprogramming (Figures 2 and 3). In their classic paper reporting the outcome of the experiment, Briggs and King (1952) showed that tadpoles developed from some enucleated eggs injected singly with blastula nuclei. At this stage, the pioneers terminated the experiment because they were interested not in cloning animals per se but rather in investigating nuclear potential. Later, however, when they tested early gastrula nuclei (Briggs and King 1960), they did rear tadpoles and showed that the majority metamorphosed into normal juvenile frogs.
Briggs and King chose donor cells from blastula and early gastrula stages because previous studies (reviewed in Spemann 1938) had shown that when regions of embryos at these stages were grafted to other areas of autologous embryos, the grafts developed according to the new site; hence, the cells of the transplanted regions were still undetermined.
If the procedure had not produced normal animals from nuclei of undetermined regions, it could not have been applied to nuclei from determined and differentiated regions to study nuclear differentiation. The totipotency of blastula nuclei was eventually demonstrated when blastula nuclei were found to direct the development of adult frogs that produced normal progeny (Xenopus laevis, Gurdon 1961; R. Pipiens, McKinnell 1962). In both studies, the donor nuclei carried a genetic marker (single nucleolus in Xenopus and pigment pattern in Rand).
Amphibian cloning:
Early gastrula cells that are fated to give rise to neural plate if left in place will, if grafted to a site destined to give rise to epidermis, develop as epidermis. On the other hand, late gastrula cells that are destined to give rise to neural plate differentiate into neural cells when placed in anatomically inappropriate sites. Thus, late gastrula cells, although not yet differentiated as neural cells, are determined to follow a neural pathway (Spemann 1938).
The transfer of embryonic nuclei:
King and Briggs (1956) found that with increasing embryonic age, more and more transplanted nuclei displayed a loss of differentiation potential. This observation was confirmed by other investigators for several frog and salamander species (reviewed by McKinnell 1978, Di Berardino 1997a, 1997b). What made this observation particularly compelling was the fact that similar results were obtained by King and Briggs (1956) with R. Pipiens and by Gurdon (1974) with the South African clawed toad, X. Laevistwo very different anuran species. X. Laevis is primitive and aquatic, and it develops rapidly, whereas R. Pipiens is more evolutionary advanced and terrestrial much of the year, and it develops more slowly Yet despite the evolutionary differences between the animals, similar loss of nuclear potential was observed in the transplanted nuclei from both species (reviewed by McKinnell 1972).
Although endoderm cells of tailbud-stage R. Pipiens embryos have lost nuclear potential, Hennen (1970) was able to reverse these otherwise stable nuclear changes with technical modifications to the transplantation procedure. She added the polycationic amine, spermine, to complex chromatin proteins and lowered the environmental temperature to lengthen the cell cycle of the oocyte host. Of the complete blastulae that developed from enucleated egg cells into which tailbud-stage nuclei had been transplanted using these modifications, 62% developed into normal larvae. By contrast, when the modifications were not used, only 25% of the blastulae develops into normal larvae. Almost 30 years ago, Hennen (1970) concluded that if normal differentiation involves the selective repression of genetic information, then repression, however stable it might be under normal conditions, is reversible as far as nuclei from tailbud presumptive midgut are concerned. The implication of Hen-nen's work is that modifications in the nuclear transfer procedure can enhance the developmental expression of nuclei from advanced developmental stages.
Cloning of larval Ana adult nuclei:
Only a small minority of larval amphibian nuclei directed enucleated eggs to develop into fertile frogs: one gut nucleus from Pleurodeles prehatching larva (Aimar 1972) and, from Xenopus swimming larvae, 20 gut nuclei (Gurdon 1962), 2 intestinal nuclei (Gurdon and Uehlinger 1966), and 2 epidermal nuclei (Brun and Kobel 1972, Kobel et al. 1973). However, it is not known whether the few totipotent nuclei originated from differentiated cells or from contaminating stem cells. In contrast to the few totipotent amphibian nuclei from larval stages, no adult nuclei were found to be totipotent (reviewed by Di Berardino 1997a, 1997b).
Extensive nuclear transfer studies of differentiated larval and adult cells from R. Pipiens and X. Faevis revealed the nuclei of these cells to be multi-potent but not totipotent: Xenopus nuclei from melanophores, erythroblasts, skin, and lymphocytes injected into enucleated eggs directed the development of pre- or post-hatching tadpoles (reviewed by Di Berardino 1997a, 1997b). The most advanced tadpoles ensued from R. Pipiens erythrocyte nuclei from juvenile frogs: 7.8% directed the formation of feeding tadpoles that survived for up to a month (Di Berardino et al. 1986).
These studies are important for several reasons. One is that the terminally differentiated state was obvious by the shape and color of the donor red blood cell. The cell type of donor nuclei in a cloning experiment is not always known because of the complexity of biological tissues that are composed of both differentiated and stem cells. With mature erythrocytes, there can be no doubt as to the cell type because of their oval morphology and red color. Furthermore, the unique morphology of the mature amphibian nucleated blood cell is always associated with a particular phase of the cell cycle (G0, in this instance) and the virtual absence of transcriptional activity.
Another notable aspect of the study was that erythrocyte nuclei were first injected into metaphase I oocytes and conditioned for one day while the oocytes matured into metaphase II oocytes (the conventional host). The matured oocytes were then parthenogenetically activated by insertion of a glass needle, and the maternal (oocyte) nucleus was removed, leaving only the erythrocyte nucleus in the cytoplasm.
On the following day, blastulae had developed that then became nuclear donors for enucleated metaphase II oocytes. These cloned erythrocyte embryos developed into feeding larvae with hind limb buds (Figure 4). The use of metaphase I oocytes for conditioning the erythrocyte genome was based on previous studies, in which erythrocyte nuclei transferred to metaphase II oocytes failed to promote development of the host beyond the early gastrula stage; however, those nuclei exposed first to metaphase I and then metaphase II oocyte cytoplasm directed the hosts to develop into larvae (Di Berardino and Hoffner 1983).
These experiments were based on the hypothesis that molecular components of the oocyte cytoplasm that prepare oocyte chromosomes to participate in fertilization would similarly condition the genetic material of erythrocytes. Obviously, the developmental potential of these terminally differentiated erythrocyte nuclei was enhanced by this conditioning; however, the actual molecular mechanisms responsible for this nuclear reprogramming require investigation.
Finally, the experiments resulted in the most develop-mentally advanced cloned animals produced with adult nuclei before the advent of Dolly, the sheep cloned from an adult mammary gland cell by Wilmut et al. (1997). Like the larvae in the erythrocyte study, Dolly developed from a quiescent nucleus, one in the G0 part of the cell cycle. One cannot but ponder the significance of the quiescent phase associated with the successful results of these donor nuclei. Perhaps the successful results of these donor nuclei in the quiescent phase suggest that G0 nuclei integrate more normally into the cell cycle of the host than those in the other cell cycle phases (Campbell 1999).
Mammalian cloning:
Cloning of mammals via nuclear transfer was initially reported in mice during the early 1980s, approximately 30 years after the first tadpole clones were produced. Although there was considerable interest in extending cloning to mammalian species, these efforts were delayed until numerous technical parameters were modified for the small (approximately 100 µm in diameter) mammalian oocyte, including enucleation and in vitro culture of viviparous oocytes and embryos.
Investigators using mammalian cells were interested in the same fundamental question of nuclear potency during embryogenesis that was posed for cells of invertebrate and vertebrate species. In fact, a series of experiments involving blastomere separation and fusion, bisection of blastocysts, and injection of inner cell mass cells into the blastocyst had already demonstrated that totipotency is maintained in mammals until at least the blastocyst stage (reviewed in Di Berardino 1997a, 1997b). In addition to these basic studies, investigators seek to improve the genetic content of livestock species to benefit agriculture focused on nuclear transfer to oocytes of large domestic animals.
Nuclear transfers in mammals are performed primarily by cell fusion, using a procedure similar to the one originally developed for murine nuclear transplants (McGrath and Solter 1983) but with some modifications (Figure 5). A micropipette is inserted through the zona pellucida (i.e., the noncellular envelope surrounding the mammalian oocyte) and positioned over the oocyte's spindle; the micropipette is then used to withdraw the spindle, chromosomes, and first polar body.
Next, a micropipette containing an intact, genetically marked donor cell is inserted through the zona pellucida, and the cell is gently injected into the cavity between the oocyte's membrane and the zona pellucida. The two cells are usually fused by electro-fusion, whereby the electrical discharge causes breaks in the cell membranes of the oocyte and donor cell, permitting the contents of each to mingle before membrane healing. The electrical discharge can activate some oocytes, although in most cases, additional treatment is required.
Alternatively, in some mouse experiments, nuclei may be injected into oocytes that are subsequently activated (Wakayama et al. 1998, Wakayama and Yanagimachi 1999). The nuclear transplants are reared in vitro through various cleavage stages and then transferred to the uteri of surrogate mothers. Totipotency has been demonstrated for nuclei from various preimplantation stages of mouse and cattle (reviewed by Sun and Moor 1995, Di Berardino 1997a, 1997b, Fulka et al. 1998). In all cases, the clones developed into adults and produced normal progeny. So far, the only primates cloned by nuclear transfer are two rhesus monkeys (Figure 5) from 8-cell embryos (Meng et al. 1997). We await a report on tests of their fertility.
In some cases, multiple clones were produced from a single donor animal; these will be valuable for testing pharmaceuticals on animals with an identical genetic (nuclear) background. For example, isogenic groups were reported in murine twins and triplets (Kono et al. 1991) and in a group of over 30 animals produced by serial cloning (i.e., the cloning of clones; Park et al. 1993, Wakayama et al. 1998). Similarly, multiple calves ensued from one donor (Bondioli et al. 1990, Willadsen et al. 1991, Chesne et al. 1993), and 10 calves were produced by serial cloning (Stice and Keefer 1993). Finally, serial cloning of goat nuclei from the 32-cell stage resulted in 45 kids (Zang and Li 1998). Three identical transgenic goats were cloned from fetal cells bearing the human gene for antithrombin III, and one goat is producing the protein in her milk (Baguisi et al. 1999).
Although cloning of preimplantation mammalian nuclei has been relatively successful, the cloning efficiency from more advanced donor stages, as in amphibians, decreases. Nevertheless, a cloning efficiency adequate for commercial use has been achieved with fetal lamb and calf fibroblast and muscle cells, as well as with adult sheep mammary gland, mouse cumulus, and calf cumulus and oviductal cells.
The percentages of newborns, based on the number of embryo clones transferred to uteri, are 5-20% for lambs (7 total) from fetal fibroblast cells (Schnieke et al. 1997), 11% for calves (3 total) from fetal fibroblast cells (Cibelli et al. 1998), and 7% for calves (2 total) from fetal muscle cells (Vignon et al. 1998). With respect to the cloning efficiency from adult cells, the percentages are 3% for lambs (1 total) from mammary epithelium (Wilmut et al. 1997), 3% for mice (33 total) from cumulus cells (Wakayama et al. 1998), 1% for mice (3 total) from tail-tip cells (Wakayama and Yanagimachi 1999), 80% for calves (8 total) from cumulus and oviductal cells (Kato et al. 1998), and 10% for calves (10 total) from mural granulosa cells (Wells et al. 1999).
However, for those persons speculating on the application of cloning to humans, we emphasize the high mortality rate occurring throughout the experimental procedure. The Dolly experiment, as we will discuss subsequently, began with 434 attempts to fuse a mammary gland cell to an oocyte; the development of this one ewe represents a success rate of only 0.2%, and the remaining attempts resulted in death, either during fusion or during various pre- and postnatal stages. Although the success rates for mouse (1-2%) and calf (3.2%) cloning from adult cells are higher than for humans, the resulting mortality rate is still high.
Several studies in sheep and cattle deserve further discussion. Sims and First (1993) initially cloned four calves from cultured cells of the inner cell mass of blastocysts. This result established the feasibility of cloning from cultured cells, and it suggested that transgenic clones might be produced by transfecting cultured cells with a foreign gene and then using these cells as donor cells for nuclear transfer. Indeed, two laboratories have now produced transgenic cloned animals. Schnieke et al. (1997) cloned three transgenic lambs (two of which survived) containing the human gene for clotting factor IX. Fetal lamb cells in culture grew as fibroblasts and were transfected with constructs composed of the coding sequences for neomycin resistance and human clotting factor IX placed downstream of the promoter sequence for ovine ß-lactoglobin. The cultures were then exposed to G418, which kills all cells except those expressing neomycin resistance. Only those cells that had integrated the genes for neomycin resistance and, therefore, clotting factor IX survived and were used for nuclear transfer. The promoter sequence of the ovine ß-lactoglobin gene directs the gene for human clotting factor IX to be expressed in a tissue-specific manner, that is, in the mammary gland. After appropriate clinical trials, the protein harvested from the milk will be used to treat hemophiliacs (Wilmut 1998).
Recently, three transgenic calves were cloned from cells of a fetal fibroblastic cell line that contained a ß-galactosidase-neomycin resistance fusion gene driven by a constitutive promoter (Cibelli et al. 1998). In the future, transgenic clones can be designed to produce a variety of complex human proteins for human use. It should be noted that the production of transgenic animals by cloning is considerably more efficient than by gene injection into the pronuclei of fertilized eggs. There are several reasons for this increased efficiency, including the fact that transgenic cells can be selected in culture before nuclear transfer.
Previous Section Next Section:
Other biological problems examined by cloning:
Cloning experiments have provided valuable insight into a number of important cellular processes, such as nuclear reprogramming, cellular aging, and genomic imprinting.
Nuclear reprogramming:
The phrase nuclear reprogramming was used in frog cloning to designate the morphological and molecular changes occurring in nuclei transplanted into oocyte cytoplasm. The cytoplasm of activated eggs induces transplanted nuclei to cease RNA synthesis and synthesize DNA. The transplanted nuclei resume RNA synthesis at later embryonic stages, at the same time as embryonic nuclei from fertilized eggs begin to synthesize RNAs.
This reversibility of nuclear function also applies to specific genes (reviewed in Gurdon 1974). Moreover, during the first cell cycle of frog nuclear transplants, non-histone proteins move bidirectionally between the transplanted nucleus and the egg cytoplasm, whereas histone proteins primarily move from the cytoplasm into the nucleus (Di Berardino and Hoffner 1975, Hoffner and Di Berardino 1977). This result suggests that the chromatin proteins are being modified.
Today, techniques are available to analyze the remodeling of chromatin proteins directly. When transcriptionally inactive sperm nuclei were incubated in extracts from activated amphibian eggs, sperm-specific histone proteins were replaced by somatic histones H2A and H2B via the molecular chaperon nucleoplasmin (Katagiri and Ohsumi 1994). Similarly, erythrocyte chromatin was remodeled when the nuclei were incubated in extracts of activated amphibian eggs: somatic histones HI and Hl° were released from chromatin into the egg cytoplasm, oocyte-specific linker histone B4 and HMG1 were incorporated into remodeled chromatin, and somatic histones H2A and H4 were phosphorylated (Dimitrov and Wolffe 1996).
With respect to mammalian nuclei, recent studies in nuclear transplant embryos of mice, rabbit, pig, and cattle confirmed that changes similar to those observed in amphibian nuclear transplants also occur during nuclear reprogramming (Fulka et al. 1998). The importance of nuclear reprogramming is emphasized by the fact that incomplete nuclear reprogramming or its failure in amphibian and mammalian nuclear transplants causes abnormal and arrested development (reviewed in Di Berardino 1997a).
Scientists are only beginning to understand the molecular changes involved in nuclear reprogramming, yet this line of basic research may result in some of the most beneficial applications of cloning to humans. For example, if scientists could explain in molecular terms how a differentiated nucleus is de-differentiated, it might be possible to repair certain diseased tissuesa small amount of normal tissue could be removed from a patient and de-differentiated in culture.
After the cell population is expanded, appropriate inducers could be added to promote a desired type of cell differentiation (e.g., bone, cartilage, or muscle). Then, the tissue could be grafted to the patient's diseased areas, where the cells would be recognized as self and not rejected.
Cellular aging.
Cloning experiments have examined the replication potential of a genome during cellular aging. Normal cells cultured in vitro have a finite replication limit (Hayflick and Moorhead 1961). However, even after serial cloning through 145 cell cycles, nuclei of blastula cells were still able to direct tadpole development (Robert G. McKinnell, unpublished study). After serial transplantation to oocytes, even terminally differentiated erythrocyte nuclei that had gone through more than 110 cell cycles had the competence to direct the formation of tadpoles (Hoffner Orr et al. 1986). It is likely that somatic nuclei are rejuvenated to some large extent in oocytes because oocytes contain a large store of molecular substances that support nuclear replication and mitosis.
Further studies may reveal the mechanism of cellular rejuvenation. Telomeres, which are normally reduced in length during the aging process (Greider and Blackburn 1996, Shay 1997), may also be shortened in animal clones. Dolly and two other cloned sheep were reported to have telomeres that were shorter than those of age-matched controls (Shiels et al. 1999). Despite their shortened telomeres, the cloned sheep were vigorous and healthy. It remains to be seen if the reduced telomere lengths will have an effect during the lifetime of the sheep.
Genomic imprinting.
Nuclear transfer and molecular studies in mice elucidated genomic imprinting, a genetic mechanism that controls the differential expression of certain pairs of autosomal alleles. Mouse nuclear transplants constructed of two maternal or two paternal pronuclei fail to develop; only nuclear transplants comprising biparental nuclei form normal offspring (McGrath and Softer 1984, Surani et al. 1984). Molecular analyses of mouse embryos revealed that certain autosomal genes are differentially expressed from maternal and paternal genomes at specific times in development (Latham 1995).
Thus, an embryo with two inactive maternal (or paternal) genes fails to transcribe a necessary gene product. It is for this reason that the mammalian embryo requires a set of genes from both the father and the mother for normal development. The experimental results in mice led to the clarification of the basis of several inherited human disorders, such as Prader Willi and Angelman syndromes, which present different phenotypes but are both due to deletions in different homologues of chromosome 15: paternal deletions in Prader Willi patients and maternal deletions in Angelman patients (Driscoll 1994).
Previous SectionNext Section
Epilogue
Because of their fundamental nature, scientific discoveries in the basic sciences (e.g., anesthesia, atomic energy, recombinant DNA) occasionally lead to unanticipated deleterious applications. Knowledge in itself is amoral, but the choices for its applications reside in the ethical decisions of humans. Cloning, like other developments in basic science, was initiated to seek new fundamental knowledge. In addition to yielding information about the role of the nucleus during cell differentiation, the procedure also provided insight into basic aspects of modifying the cancer phenotype, rejuvenating aged nuclei by oocyte cytoplasm, nuclear reprogramming, and genomic imprinting. It will continue to yield new knowledge in these and other basic subjects.
As scientists who have worked in frog cloning, we are gratified to see decades of basic research culminating in the production of cloned mammals that will produce human proteins for the alleviation of human disease. Cloning will likely result in the genetic improvement of livestock, the production of animal models to study and treat human diseases, and sources of animal tissues and organs for xenotransplantation to humans. We consider these applications of cloning appropriate. Indeed, they are the reasons why Dolly was produced.
Once Dolly appeared, however, the news media became consumed with the idea of cloning human adults, which immediately stimulated a worldwide debate among ethicists, theologians, clerics, lawyers, legislators, and, of course, laypeople. We do not intend to summarize the many proposals and convictions of others; rather, we delineate briefly the reasons for our belief that human cloning is scientifically and ethically unsound. We define human cloning as the attempt to produce a human organism by any cloning procedure: blastomere isolation, bisection (splitting) of preimplantation embryos, and nuclear transplantation.
Nuclear transplantation of embryonic, fetal, or adult cells from all species results in abnormal animals at a frequency that increases with the age of the donor cell. The failures result from incomplete nuclear reprogramming and failed cell cycle matching, although other causes may in some cases be responsible. Abnormal nuclear transplants from all donor stages arrest development at various stages: nuclear transfer and activation, cleavage, organo-genesis, tadpole, and juvenile. In viviparous species, abnormal transplant embryos may fail to implant, and those that do implant may abort at various embryonic and fetal stages. Finally, those nuclear transplants that are born may die soon after birth or survive with birth defects.
Analyses of abnormal nuclear transplant embryos from frogs, mice, cattle, rabbits, and pigs revealed chromosomal and/or molecular abnormalities that could account for the morphological aberrations (reviewed in Di Berardino 1997a). Furthermore, even some normal nuclear transplant frog blastulae derived from embryonic or adult nuclei contained abnormal chromosomes in some of their cells, whereas the chromosomes in other cells of the same blastulae appeared normal. Such cases would pose confusion in monitoring normal-appearing nuclear transplants for further culture and development. For these reasons, we consider the use of any cell type (not just those from adults) for human cloning scientifically and ethically unsound.
In the case of Dolly, she was the only successful case out of 434 attempted fusions of oocytes and donor cells that were taken from cultures of mammary gland. Even as cloning of adult nuclei becomes much more efficient, there will still be hazards to humans. For example, the donor cells could suffer mutations in situ from radiation, chemicals, and/or aging during the lifetime of the donor. Mutations could also arise in the donor cells during cell culture, an event that is not unusual. There are still other scientific concerns.
Will telomere shortening in the donor cell limit the life span of the clone? Will stored gene products (RNAs and proteins) in oocytes from foreign donors always be compatible with the donor nucleus? Finally, it is important to consider that meiosis, which precedes sexual reproduction, affords humans another opportunity for DNA repair and therefore should not be avoided in favor of asexual (somatic) reproduction. For all of these reasons, we oppose human cloning.
Apollyon the Greek Sun god who was earlier worshipped during the Egyptian Eighteenth Dynasty of Akhenaten, the father of Tutankhamen, rises up on the earth in pursuit of global dominance and aggressively works to seize control over a final age of human apostasy. The Bastard child has grown up and serves in global prominence, waiting in the wings" for his reveal. Believers who pay attention to biblical prophecy know his identity and restrain him until they are taken out of the way... NOT by his hand but by the Hand of the Almighty One!
In 1907, the actual body of Akhenaten was discovered in Egypts Valley of the Kings by British archaeologist named Edward Ayrton. After unearthing Akhenatens mummified remains, Edward Ayrton was able to confirm that the ancient pharaohs skull was misshapen and elongated. Akhenaten was succeeded by his son, Tutankhamun, who became the most renowned pharaoh of all time. When his tomb was discovered in 1922 by Howard Carter, Tutankhamun was also found to have an elongated skull. In February 2010, the results of DNA tests confirmed that he was the son of Akhenaten. In 2012, , the tomb KV55 mummy was proven through DNA to be that of Akhenaten.
Amenhotep IV/Akhenaten - Birth name: Amenhotep ("Amun is satisfied"), Adopted name: (CHANGED HIS NAME) to Akhenaten/Akhenaton ("Servant of the Aten"), Throne name: Neferkheperure ("Beautiful are the Manifestations of Re"), Rule: 1350 - 1334 BC (10th king of the 18th dynasty, New Kingdom), Noteworthy relatives: Nefertiti (queen), Tutankhamun (son), Ankhesenamun (daughter), Merytaten (daughter), Smenkhkare (son), Amenhotep III (father), Tiy (mother).
He was born to Amenhotep III and his Chief Queen Tiyee and was their younger son. Akhenaten was not originally designated as the successor to the throne until the untimely death of his older brother, Thutmose. Amenhotep IV succeeded his father after Amenhotep III's death at the end of his 38-year reign, possibly after a short coregency lasting between either 1 to 2 years.
Pharaoh Akhenaten was known as the (RENEGADE), Heretic King. He was the tenth King of the 18th Dynasty. Egyptologists are still tying to figure out what actually happened during his lifetime as much of the truth was buried after he died.
Akhenaten lived at the peak of Egypt's imperial glory. Egypt had never been richer, more powerful, or more secure. Up and down the Nile, workers built hundreds of temples to pay homage to the Gods. They believed that if the Gods were pleased, Egypt would prosper. And so it did.
A religious reformer he made the Aten, the sun disc, the center of Egypts
Gem-pa-Aten temple at East Karnak - "the Aten is found" - Cairo Museum
O sole god without equal !
You are alone, shining in your form of the living Aten.
Risen, radiant, distant and near.
Great Hymn, 47 & 73-74.
SOURCES: Egyptologist Jan Assmann
Oxford Journals: http://bioscience.oxfordjournals.org/content/49/11/875.full
University of Maryland Medical Center: Akhenaten Deformities | University of Maryland Medical Center http://umm.edu/ /a-feminine-physique-a-long-thin-neck-and-e
http://umm.edu/ /a-feminine-physique-a-long-thin-neck-and-e
1 Professor of Genetics and Cell Biology at the University of Minnesota, Saint Paul, MN 55108-1095. He is the author of Cloning: Nuclear Transplantation in Amphibia (1978) and Cloning of Frogs, Mice and Other Animals (1985) and the 1998 recipient of the Prince Hitachi Prize in Comparative Oncology, awarded by the Japanese Foundation for Cancer Research. E-mail: mckinnel@biosci.umn.edu
2 Professor emerita of Biochemistry at MCP Hahnemann University, Philadelphia, PA 19129. She is the author of Genomic Potential of Differentiated Cells (1997) and the 1996 recipient of the Jean Brachet Memorial Award, given by the International Society of Differentiation, Inc..
http://beforeitsnews.com/ /youre-traveling-through-another-
http://www.livescience.com/39349-akhenaten.html
http://bioscience.oxfordjournals.org/content/49/11/875.full
http://conservativetribune.com/carson-criticizes-obama-se /
http://www.neferchichi.com/akhenaten.html
http://www.youtube.com/watch
http://www.heptune.com/Marfans.html
http://www.ancient-code.com/akhenaten-the-alien-pharaoh/
World leaders will announce that "Hyper Intelligent Alien" biological founders of the human race have provided indisputable evidence that proves that the origin of human development is the product of the ancestral inception of Alien male DNA. The Anti-Christ' father is the devil, he is called the "Son of Perdition", the offspring of "the father of ALL Lies"
Ancient Biblical history records the existence of "other beings" and their presence and sphere of influence throughout history on planet Earth. THE "BIG LIE" is that these entities are not "ALIEN" Hybrids from another planet but rather ANGELS, (fallen angels), who were expelled from the presence of God in ancient times. These creatures disguise themselves at "beings of light". Isaiah 14:12 / Luke 10:18 / Revelation 12:4.
Biblical History records that in ancient times, these "fallen beings" bred with some human women, creating a race of giants known as Nephilim. Nephilim embodied corrupt DNA and subsequently attempted at various intervals throughout human history, to physically dominate the Earth and its inhabitants. Both the fallen angels and their offspring the Nephilim, have wreaked havoc on earth every since the corruption of Adam. Read more:
Adam's corruption ran deep into his DNA. When Adam yielded himself to the Serpent, (Lucifer / Satan), who is the "commander and chief" of these "beings", the portal of another dimension, (Hell), literally opened onto the Earth and corruption entered into Adam and damaged the precious, Divinely designed and exclusive gift of incorruptible human DNA that held the unique genetic properties, "strands and markers", of life eternal.
Adam's broken relationship with his Creator ensured that he, Adam, would not live in his corrupted state of being forever. This is the very reason why a corrupt man needed a pure incorruptible God to intervene in this matter and provide a Divine Answer to satisfy the legal requirements rendering the eternal justice of a perfect God that would release man from the corruption of death.
Once the DNA was damaged, Satan gained access through man, to the entire Planet Earth, its world systems and future human race launching the "BIG LIE". Genesis 6:16
THE FINAL BIG LIE-
It will be announced in the near future that it was necessary to conceal the true identity of Barack Hussein Obama's "paternal" father in order to reveal to the world, at an appropriate moment, his true biological connection to ancient alien ancestry. 2 Cor 11:14. (It is the "illusion" of this claim that will make converts accept this final perverted global plot to rule the Earth.)
This announcement may occur sometime in the year 2025 and be followed by uncensored information from government sources revealing decades of "eighth kind" encounters and ongoing alien dialog with world leaders confirming this claim. It is at this time, with the assistance of Pope Francis that he will be received as a god sent to the inhabitants of Earth to save mankind.
In a final act of defiance against God, a last attempt to circumvent the Will of God and rule over planet Earth and inhabitants is now taking place in our generation.
The Truth is that God Himself became flesh, A virgin conceived the Seed of Jehovah and the Divine offspring, Messiah, Jesus The Christ was born to die in our place. The innocent for the guilty, the Lamb of God, slaughtered in the place of man, The Guilt Offering who took away the sins of all men and its sting of death for all who call upon His Name. John 3:16. HELL... THE (mAN of Lawlessness) may be the hideous creation of "ARTIFICIAL INSEMINATION" from the Ancient DNA of Akhenaten. THIS SON of PERDITION, "whose father IS the devil, carries out his father's desires. And like his father, who was a murderer from the beginning, not holding to the Truth, for there is no Truth in him. When he lies, he speaks his native language, for he is a liar like his father, the father of lies".
Hussein's fraudulent birth certificate was designed to not just cover "WHERE" he was born, but to ALSO not disclose "WHO" his genetic paternal, "father" honestly is. The "COVER-UP" will give credence to the future claim by Francis, the false prophet of Rome, that this "anti-Christ" is the "PROOF", (evidence) of the superior race of the extra-terrestrial genetic "fatherhood" an alien origin of human race.
The "man of lawlessness" is the appointed representative of an "extra-terrestrial" entity claim of human "origin" who has returned to earth to "save their own creation" from demise. As "man" will be presented to humanity, whose "paternal", (of the male genetic origin) is undocumented (not known) and whose mother, (maternal) genetic origin), is of a "whore" by nature, the exact opposite of Yeshua Messiah! This appointment by Hell will manifest at the time of the coming global government "alien among us disclosure". MOST of humanity will embrace the idea, thus Satan's GREAT DECEPTION, that will rob Heaven of scores of souls, who will follow this BEAST straight to Hell.
Terra firma (of earth) vs Extraterrestrial (Originating from outside of the Earth's atmosphere, from space, or from another planet; alien to Earth or its environment). Believers will need to cling to every Word of YHWH to overcome the lies that are manifesting about the human origin. Genesis specifically proclaims the "man" is the exclusive creation of YHWH taken from "TERRA FIRMA", (EARTH). Remember this when the proclamation is that man is from genetic alien extraterrestrial origin!!!
"the LORD God formed man out of the clay of the ground (terra firma) and blew into his nostrils the breath of life, and so man became a living being.
A "BASTARD CHILD" of HELL IS "BORN"... Born of a whore. THE (mAN of Lawlessness) may be the hideous creation of "ARTIFICIAL INSEMINATION" from the Ancient DNA of Akhenaten. THIS SON of PERDITION, "whose father IS the devil, carries out his father's desires. And like his father, who was a murderer from the beginning, not holding to the Truth, for there is no Truth in him. When he lies, he speaks his native language, for he is a liar like his father, the father of lies".
Hussein's fraudulent birth certificate was designed to not just cover "WHERE" he was born, but to ALSO not disclose "WHO" his genetic paternal, "father" honestly is. The "COVER-UP" will give credence to the future claim by Francis, the false prophet of Rome, that this "anti-Christ" is the "PROOF", (evidence) of the superior race of the extra-terrestrial genetic "fatherhood" an alien origin of human race.
The "man of lawlessness" is the appointed representative of an "extra-terrestrial" entity claim of human "origin" who has returned to earth to "save their own creation" from demise. As "man" will be presented to humanity, whose "paternal", (of the male genetic origin) is undocumented (not known) and whose mother, (maternal) genetic origin), is of a "whore" by nature, the exact opposite of Yeshua Messiah! This appointment by Hell will manifest at the time of the coming global government "alien among us disclosure". MOST of humanity will embrace the idea, thus Satan's GREAT DECEPTION, that will rob Heaven of scores of souls, who will follow this BEAST straight to Hell.
Terra firma (of earth) vs Extraterrestrial (Originating from outside of the Earth's atmosphere, from space, or from another planet; alien to Earth or its environment). Believers will need to cling to every Word of YHWH to overcome the lies that are manifesting about the human origin. Genesis specifically proclaims the "man" is the exclusive creation of YHWH taken from "TERRA FIRMA", (EARTH). Remember this when the proclamation is that man is from genetic alien extraterrestrial origin!!!
"the LORD God formed man out of the clay of the ground (terra firma) and blew into his nostrils the breath of life, and so man became a living being.
O'Akhenaten, O-Bama & Mengele:
*** Between April and May 1960, Dr. Mengele was confirmed to have been in Buenos Aires. Just before or after the capture of SS Eichmann, Mengele disappeared off the radar map. Obama II was born in August 1961. Normally, it would mean that he was conceived in December 1960 during the period of Dr. Mengeles most secretive disappearance episode during the early 1960s. In fact, his trail remained cold until he reportedly surfaced in Brazil in the late 1960s.
Born August 4, 1961, Barry Soetoro, who changed his name to Barack Hussein Obama continues to Keep his Past Identity cloaked in a Shroud of Secrecy. Dr. Ben Carson says he wants President Obama to come clean about the shroud of secrecy surrounding his past, including his birth records.
On November 24, 1982, while Obama II was a student attending Columbia University, Barack Obama, Sr. Was found dead in Kenya. The cover story is that Obama, Sr. Drove his car into a tree following a day of drinking in Nairobi. However, Obama, Sr.,s family, sister, Hawa Auma Obama, and his friend Patrick Ngei believe that he had been assassinated, Obama didnt die out of pure accident . Bright Luos were eliminated by the Kenyatta government.
So, what did Stanley Ann really mean when she suggested to her friends that she could bear children without dating and marriage? Her comment suggests an unusual acute awareness for 1960 of the existence of in vitro fertilization, embryo transfer, and artificial insemination technology- positive eugenics.
Out of everyone in the world, SS Dr. Josef Mengele and the underground SS Knights of the Black Sun had been the most secretive, treacherous and satanic forces that would have conceived such a diabolical plan.
Dr. Mengele disappeared during the time of conception and birth of Obama II. During that time, Dr. Mengele was secretly deeply involved in clandestine artificial insemination in human beings. When Obama II and Stanley Ann returned to Hawaii in 1962, Dr. Mengele was exposed by Argentine historian Jorge Camarasa artificially creating the master race of blond haired, blue eyed Aryans in twins by the dozens in Candido Godoi, Paraguay. Barry Soetoro, (Hussein Obama) was just part of the experimental process of the CIA and was raised with intent and purpose always within their grasp and oversite.
Dont forget pursuant to the SS SCIENTIFIC SATANIC PRINCIPLE of the DIVINE POWER of OPPOSITES that if they had the power to create the MASTER RACE in duality, they had to power to create the SUBSERVIENT.
They had the money, technology, resources, clandestine support, international networks and motive to put a covert Diabolical Black Human Cyborg in the Whitehouse to continue the 1000 Year THIRD REICH and its Satanic Principle of Aryan Planet Domination and the Annihilation of People of Color. An African-American Brother? You never seen anything like him. KEEP DREAMING THAT THE BOYS FROM BRAZIL IS A WORK OF PURE SCIENCE FICTION- AND THAT FAT MEAT AIN'T GREASY!
Dr. Josef Mengele and the SS developed Artificial Insemination and Fertilization that even still today THE CULT OF THE SS stand at the pinnacle and forefront of Modern Genetic and Reproductive Technology. (VN: and that alone should make you damn nervous about the future existence of the human race, if they do away with all of us and only have their clones to deal with, then they are safe and exposed and can live like animals.
Beyond Lebensborn, Himmlers Secret - CIA- All in the Family:
British Intelligence Advisor: CIA Conducted DNA Test on Obama Found No Match to Alleged Grandparents.
Stanley Anns father, Stanley Armour Dunham, had been Obama, Sr.s CIA handler. Stanley Anns mother, Madelyn Dunham, above, also managed the agencys secrets. In 1960, she started working at Hawaii's largest bank, Bank of Hawaii, and was promoted to be one of the first female bank vice presidents in 1970. Madelyn Dunhams expertise had been in the banks escrow accounts department described to be primarily used for 3rd party money transfer accounts between two people who basically do not trust each other and still need a deal to be done. Madelyn handled the secret escrow-payoff accounts of the CIA front company, Bishop Baldwin Rewald Dillingham Wong (BBRDW), Ltd.
BBRDW was primarily run by Ronald R. Rewald. Rewald was a CIA agent recruited to spy on the student radical movement in America in the 1960's. Now Bishop Baldwin expanded its operations greatly. Satellite offices opened up in more than a dozen cities worldwide. It now employed a staff of nearly 200 people. Rewald lived in a Hawaiian estate near Diamond Head valued at over a million dollars in 1980. Bishop Baldwin had a fleet of cars and a chauffeur to drive around Rewald and Bishop Baldwins clients. The company which had four accounts at its incorporation in 1979 had 110 by 1983. And in such exotic places as Hong Kong and the Cayman Islands.
Rewald was now meeting with people like the Sultan of Brunei and Vice-President George Bush and arranging secret arms deals with Rajiv Gandhi of India. In fact, this last seems to have been the major CIA use of the company I.e. To spirit weapons and arms of all types into Pacific Rim countries. Bishop Baldwin also used businessmen to collect intelligence and to direct the flow of capital into American companies. It also was used as a cover for more sinister assignments like the assassinations of leftist leaders and sympathizers.
BBRDW and Rewalds cover was blown in Hawaii by a tip to a news reporter in 1983. Just prior to their cover being blown, CIA Special Agent Jon S. Lodeesen just happened to show up at their doorstep January 12, 1983 for a special assignment in Hawaii.
During the Falkland Islands War of April 1982 between Argentina and Britain, Ron Rewald met with the Gehlen Orgs SS/CIA representatives in Uruguay, Licio Gelli of the Masonic Lodge P2, Klaus Barbie and Heinrich Rupp, to arrange for the Argentine purchase of the French-made Exocet missile used in war.
BBRDW and Rewald had also been a pipeline for the Reinhard Gehlen Org. Madelyn Dunham retired from the Bank of Hawaii in 1986, so it is highly likely that Stanley Anns mother, Madelyn Dunham, handled the Gehlen Orgs secret transfer account to buy Exocet Missiles through the Bank of Hawaii. Stanley Ann Dunham had been a multi-generational asset deeply embedded in the Reinhard Gehlen SS-CIA fraction.
The perfection of positive eugenics was a top-secret mission of the THIRD REICH and a clandestine project of Reichsfuhrer SS-I Heinrich Himmler and the SS. Even today, two of Himmlers SS doctors stand at the pentacle of Human Fertilization and Embryology to Fabricate Aryans from scientific genetic material; and in duality (Satanic Principle) Annihilate non-Aryan genetic material.
(VN: given what we have since learned, it may well be the other way around. Remember, the gov to this day is taking the DNA of newborn babies and following and tracking RH Neg citizens without any reason given. We covered much here about RH negs and they are the ones that are Aryan, and Hitler was obsessed with them and their blood negative is found nowhere in nature. Hmmm, wonder what that means? A different species?).
Himmlers top in vitro fertilization and artificial insemination specialists were SS Teutonic-Knights of the Black Sun and neither one of them were tried at Nuremberg. One was captured in 1945 by the Soviets and may have been turned into a Soviet mole in West Germany, and the other escaped justice under the shroud of the Gehlen Org and Operation CAMPUS.
In 1961-1962, this agency doctor was in Brazil with Lodeesen and Jim Jones of the Peoples Temple. This particular doctor was known to be involved in clandestine artificial insemination and in vitro fertilization projects. He is the subject of the popular 1978 movie thriller, The Boys From Brazil.
Dr. Mengele and Obama II: Timeline of the BOY FROM BRAZIL.
Jim Jones idolized Dr. Josef Mengele. There is cause to believe that while in Brazil in 1961, Jones had met his mentor and had been initiated into the CULT OF THE SS. Additionally, Jones Peoples Temple mind control experiments were an extension of Mengeles Auschwitz experiments.
At Mendocino State Mental Hospital where the entire staff was dismissed and replaced by Jim Joness personnel, according to mind-control survivors, Dr. Mengele had been there in association with Jim Jones programming patients and inmates. It is reported that Dr. Mengele also visited Jonestown, Guyana.
On May 11, 1960, the Israeli MOSSAD captured Obersturmbannführer SS Otto Adolf Eichmann, one of the major organizers of the Final Solution in San Fernando, an industrial community north of the center of Buenos Aires.
MOSSAD agents had been in Argentina since April 1960. At the time, Dr. Mengele was also discovered to have been living in Buenos Aires, Argentina. MOSSAD agents found Dr. Mengeles address and actually took pictures that they determined beyond any doubt the identity of the notorious Demon of Death of Auschwitz. The MOSSAD walked away from Dr. Mengele, one of the worlds most wanted Nazi war criminals. I gather that he was a little too big for their boots protected by U.S. National Security like SS Baron Otto von Bolschwing.
*** Between April and May 1960, Dr. Mengele was confirmed to have been in Buenos Aires. Just before or after the capture of SS Eichmann, Mengele disappeared off the radar map. Obama II was born in August 1961. Normally, it would mean that he was conceived in December 1960 during the period of Dr. Mengeles most secretive disappearance episode during the early 1960s. In fact, his trail remained cold until he reportedly surfaced in Brazil in the late 1960s. [87]
Now, you should have some idea where Stanley Ann could have got her notions of having children without natural procreation-beyond Lebensborn and why Jim Jones showed up in Hawaii. She had been primed for the job as a multi-national CIA/MK ULTRA asset by the satanic (Negative Eugenics) plans of Dr. Josef Mengele; and Jim Jones showed up in Hawaii to dust the trail of his mentor and master.
Dr. Mengeles Land of Aryan Twins:
In a recent book, Mengele: The Angel of Death in South America, the Argentine historian Jorge Camarasa, a specialist in the post-war Nazi flight to South America, contends that Dr. Mengele re-surfaced in about 1962 or early 1963 in the enclave of Colonias Unidas, Paraguay that was founded by German immigrants in 1900.
From there in 1963, Dr. Mengele began to make regular trips to another predominantly German community just over the border in Brazil the farming community of Candido Godoi. (VN: Oh, how very interesting, that coincides with the fact that Bush Jr, and Rev Moon, just bought up the top half of Paraguay and since these cretins do nothing for just one reason, I am wondering if this fraud was one of the reasons? The Cloning of a US President and the need to bury any evidence of what may well have happened there?).
In Candido Godoi, people remembered that Dr. Mengele talked of artificial insemination in human beings, and that he continued working with animals, proclaiming that he was capable of getting cows to produce male twins.
Nobody knows for sure exactly what date Mengele arrived in Candido Godoi, but the first twins were born in 1963, the year in which we first hear reports of his presence, Camarasa said. I think Candido Godoi may have been Mengeles laboratory, where he finally managed to fulfill his dreams of creating a master race of blond haired, blue eyed Aryans, he said.
Dr. Mengele also worked out of the infamous post-war compound of Cologna Dignidad near Paral, Chile founded in 1961 by Nazis straight out of Germany. The UN commission found that the above Dictator for life General AUGUSTO PINOCHETs military intelligence agency DINA-sponsored torture chambers at Colonia Dignidad.
Its underground bunker and chambers were used as research center to continue Dr. Mengeles work of refining the art of torture, prisoners have allegedly been subjected to different experiments without any interrogation Prisoners charge that torture is personalized through an initial interrogation which establishes the personal traits of the individual This data is then used to program the torture sessions so that the result is a totally debilitated person who will comply with any demand.
According to a UN report, in Colonia Dignidad there is a specially equipped underground torture center with small soundproofed cells, hermetically sealed. The detainees heads are covered with leather hoods, which are stuck to their faces with substances that are supposedly chemicals. In these cells, interrogations are carried out through electronic equipment, including loudspeakers and microphones, while detainees are tied naked to metal frames to receive electric shocks.
(VN: oh, wow, now Gitmo makes a lot more sense than it ever did before and given the results where a man was so mind controlled he agreed to blow up his own balls in his pants, and not say a word, then you know he was mind controlled. I told you these people are absolutely insane, and add in the bloodline issue, they are doubly insane. We have to shut down GITMO FOR THE SAKE OF HUMANITY. We are guilty if we let this continue in our name as the people of this country.)
The CIA Medical Experimental Compound at Jim Jones field station and agricultural commune in Jonestown, Guyana had been established as a U.S. Shadow of Cologna Dignidad. Jeannie Mills, a white overseer and ring leader of the Peoples Temple that had been assassinated in Berkeley in 1980, claimed to have seen films taken inside a Chilean torture camp while at Jonestown. These films were either taken at the Colonia Dignidad or another located at Pisagua. At that time, the only source possible for the films had to have been the CIA, DINA or Dr. Mengele.
Contrary to HollyWeird and government propaganda, Dr. Mengele didnt appear to be a frightened fugitive constantly on the run from the law. To the contrary, after the capture, conviction and execution of Obersturmbannführer SS Otto Adolf Eichmann in 1962, the Great Nazi Satanist appeared to have travelled freely throughout South America, Canada, Germany and the United States without a care in the world polarizing the populations, spreading death, destruction and CHAOS.
In 1985, Senator Alfonse M. DAmato (R-N.Y.) testified at a Senate hearing that the U.S. Government had evidence from Cables and other documents indicate that Mengele had a network available to him in raising large sums of money and that high officials (in South America) gave him protection
In other words, Dr. Mengele didnt die in 1979 and was right in the middle of the CIA Iran-Contra CRACK Cocaine Scandal. Of course, U.S. Government agencies claimed they didnt know what Senator DAmato was talking about, because the CIA had also been protecting and supporting him since it was formed in 1947.
Out of everyone in the world, SS Dr. Josef Mengele and the underground SS Knights of the Black Sun had been the most secretive, treacherous and satanic forces that would have conceived such a diabolical plan.
Dr. Mengele disappeared during the time of conception and birth of Obama II. During that time, Dr. Mengele was secretly deeply involved in clandestine artificial insemination in human beings. When Obama II and Stanley Ann returned to Hawaii in 1962, Dr. Mengele was exposed by Argentine historian Jorge Camarasa artificially creating the master race of blond haired, blue eyed Aryans in twins by the dozens in Candido Godoi, Paraguay.
Dont forget pursuant to the SS SCIENTIFIC SATANIC PRINCIPLE of the DIVINE POWER of OPPOSITES that if they had the power to create the MASTER RACE in duality, they had to power to create the SUBSERVIENT.
They had the money, technology, resources, clandestine support, international networks and motive to put a covert Diabolical Black Human Cyborg in the Whitehouse to continue the 1000 Year THIRD REICH and its Satanic Principle of Aryan Planet Domination and the Annihilation of People of Color. An African-American Brother? You never seen anything like him. KEEP DREAMING THAT THE BOYS FROM BRAZIL IS A WORK OF PURE SCIENCE FICTION- AND THAT FAT MEAT AIN'T GREASY!
In Part II, I intend to show where, when and how Dr. Josef Mengele and the SS developed Artificial Insemination and Fertilization that even still today THE CULT OF THE SS stand at the pinnacle and forefront of Modern Genetic and Reproductive Technology. (VN: and that alone should make you damn nervous about the future existence of the human race, if they do away with all of us and only have their clones to deal with, then they are safe and exposed and can live like animals.
The resulting frogs expressed the donor genetic markers, thus proving that the frogs were derived from donor nuclei and not oocyte nuclei left behind from faulty enucleation.
Many other investigators in the United States, England, France, China, and Japan soon entered the field. They too confirmed the totipotency of blastula nuclei in other frog species as well as in some salamander species (reviewed in Di Berardino 1997a).
The pioneers of nuclear transplantation concluded in a classic understatement that although the method of nuclear transplantation should be valuable principally for the study of nuclear differentiation, it may also have other uses (Briggs and King 1952, p. 462). We discuss this research and other applications of cloning below. In retrospect, it is clear that the amphibian nuclear transplantation procedure became the prototype for cloning multicellular animals.
Although the method of nuclear transplantation should be valuable principally for the study of nuclear differentiation, it may also have other uses. (Briggs and King 1952, p. 462)
Clearly, 1997 was the year of the clone (Figure 1). The cover illustration of Nature (27 February 1997) announced the birth of Dolly, the ewe cloned from an adult sheep in Scotland, and Science (19 December 1997) proclaimed Dolly to be the breakthrough of the year. Newspapers, news magazines, radio, and television were even more fervent in their reports of Dolly.
Even now, several years later, Dolly (Wilmut et al. 1997) and a number of mice (Wakayama et al. 1998, Wakayama and Yanagimachi 1999) and calves (Kato et al. 1998, Wells et al. 1999), all cloned from adult cells, continue to evoke fascination. Was Dolly the first animal to be cloned? Of course not. Why, then, the sudden, almost unprecedented attention to the announcement of a cloned sheep? We believe that because Dolly was the first animal cloned from an adult cell, she stimulated scientists, theologians, ethicists, journalists, and politicians to contemplate the application of cloning to humans.
The point of this article is not to reconsider the extension of cloning to humans, a subject that has already been covered (e.g., Silver 1997, Kolata 1998, Nussbaum and Sunstein 1998), but to consider the genuine rationales that stimulated the original and continuing efforts in cloning research. Cloning was never intended as a procedure for the simple multiplication of animals.
Frogs are cheap in the United States, as are sheep in Scotland. The reasons for cloning are more complex than simply producing identical animals, and in this article, we consider those reasons and the results that have been obtained with the procedure. We place cloning in the historical context of developmental biology and review results obtained with the procedure. Some readers may wonder how two scientists with cloning experience view the ethics of human cloning. We offer our views in the epilogue.
Cloning's roots in nineteenth-century biology:
How do cells become specialized during development? One cell, the zygote, gives rise to a multiplicity of cells that in time become increasingly specialized, or differentiated. In the latter part of the nineteenth century, August Weismann believed that differentiation results from the differential and sequential partitioning of the genome as the cells divide (reviewed by Wilson 1928, Spemann 1938).
The attractiveness of the now-discarded Weismann hypothesis was that it could be tested. Wilhelm Roux, in an 1888 experiment (Spemann 1938), killed one cell (blastomere) of a two-cell amphibian embryo and found that a half-embryo developed, suggesting that some genes are lost during cell replication. However, in 1892, Hans Driesch found that if the blastomeres of two-cell sea urchin embryos were physically separated, entire embryos formed from each blastomere (Spemann 1938); similar results were obtained when amphibian blastomeres were isolated, provided that they contained a portion of cytoplasm known as gray crescent material. Thus, the genome was not diminished, but rather reproduced during cell division. (Roux's half-embryos were later interpreted to result from the inhibitory effect of the dead blastomere.)
CLONING HAS PROVIDED INSIGHTS INTO NUCLEAR DIFFERENTIATION, NUCLEAR REPROGRAMMING, CELLULAR AGING, AND GENOMIC IMPRINTING:
Blastomere separation of embryos beyond the 2- to 16-cell stage (depending on species) was noninformative for testing genomic potential because the cells had too little cytoplasm. But in 1894, Jacques Loeb fortuitously observed that fertilized sea urchin eggs sometimes ruptured when exposed to hypotonic solutions (Spemann 1938).
The extruded portion of the egg was usually bereft of a nucleus and remained uncleaved. Occasionally, nuclei traversed the isthmus between the cleaving egg and the extruded cytoplasmic material. In such cases, the extruded cytoplasm cleaved along with the main mass, resulting in the formation of two entire embryos. In 1914, Hans Spemann performed a conceptually identical experiment on an amphibian egg (Spemann 1938).
He constructed a zygote with a noose made of baby hair, causing the egg to assume the shape of a dumbbell. When the cleaving (nucleated) portion reached the 8- or 16-cell stage, he loosened the constriction and permitted a nucleus to move to the non-nucleated cytoplasmic portion. Here, too, the non-nucleated portion cleaved and formed a clone of its nuclear donor.
These primitive nuclear transplantation (cloning) experiments affirmed the view that the complete genome is replicated during cell division, at least during early cleavage. The stage was now set for modern cloning experiments to examine the genomic potential of older embryonic cells. However, Spemann (1938) could see no way for the moment to manually insert a nucleus from older embryos into enucleated cytoplasm. Robert Briggs and Thomas J. King found a way.
Success in Philadelphia:
Around 1943, Briggs (1911-1983), an embryologist working at what is now the Fox Chase Cancer Center in Philadelphia, wanted to determine whether the genome of older somatic nuclei remains equivalent to the zygote nucleus throughout development (Patterson n.d.). One possible approach was to transplant a somatic nucleus into an oocyte whose own nucleus had been removed and then observe what type of development occurred.
Briggs was aware that Comandon and de Fonbrune (1939) had transplanted single nuclei in the unicellular amoeba, but he recognized that nuclear transfer in the oocytes of metazoans would be more arduous because of their complex physiological and biochemical requirements for embryonic development.
Over the next 7 years, while analyzing the development of haploid and triploid frog embryos, he became experienced with various microsurgical techniques that would contribute to the procedure of nuclear transfer. In 1948, one of us (Di Berardino) joined Briggs's laboratory and was there to witness the first cloning of metazoan animals in 1952.
In 1949, Briggs began searching for a research fellow to develop a nuclear transplantation procedure for the North American leopard frog, Rana pipiens. He also applied to the National Institutes of Health for funds to support the project. His first attempt was rejected because the reviewers considered his proposed research a harebrained scheme with little chance of success
However, a second application was successful, and Briggs brought King in as his research fellow in 1950. The two scientists first determined that eggs lacking a functional nucleus but containing a cell division center developed at best into partial blastulae (Briggs et al. 1951). This baseline study established the total developmental potential of the recipient host; any further development could, therefore, be credited to an introduced nucleus.
Briggs and King's nuclear transfer procedure involved first activating an oocyte at metaphase II of meiosis by pricking it with a glass needle, which initiated the metabolic changes normally induced by the sperm. Approximately 15 minutes later, a pit (black dot) appearing on the surface of the oocyte was micro surgically removed with another glass needle; this surgery removed the oocyte's chromosomes, resulting in an enucleated egg. Finally, a blastula cell was aspirated into a glass micropipette whose lumen was slightly smaller than the diameter of the cell to gently break the cell membrane.
The broken cell was then transferred to the animal hemisphere of the enucleated egg, which permitted the nucleus to interact with the cytoplasmic molecular milieu of the oocyte and to undergo nuclear reprogramming (Figures 2 and 3). In their classic paper reporting the outcome of the experiment, Briggs and King (1952) showed that tadpoles developed from some enucleated eggs injected singly with blastula nuclei. At this stage, the pioneers terminated the experiment because they were interested not in cloning animals per se but rather in investigating nuclear potential. Later, however, when they tested early gastrula nuclei (Briggs and King 1960), they did rear tadpoles and showed that the majority metamorphosed into normal juvenile frogs.
Briggs and King chose donor cells from blastula and early gastrula stages because previous studies (reviewed in Spemann 1938) had shown that when regions of embryos at these stages were grafted to other areas of autologous embryos, the grafts developed according to the new site; hence, the cells of the transplanted regions were still undetermined.
If the procedure had not produced normal animals from nuclei of undetermined regions, it could not have been applied to nuclei from determined and differentiated regions to study nuclear differentiation. The totipotency of blastula nuclei was eventually demonstrated when blastula nuclei were found to direct the development of adult frogs that produced normal progeny (Xenopus laevis, Gurdon 1961; R. Pipiens, McKinnell 1962). In both studies, the donor nuclei carried a genetic marker (single nucleolus in Xenopus and pigment pattern in Rand).
Amphibian cloning:
Early gastrula cells that are fated to give rise to neural plate if left in place will, if grafted to a site destined to give rise to epidermis, develop as epidermis. On the other hand, late gastrula cells that are destined to give rise to neural plate differentiate into neural cells when placed in anatomically inappropriate sites. Thus, late gastrula cells, although not yet differentiated as neural cells, are determined to follow a neural pathway (Spemann 1938).
The transfer of embryonic nuclei:
King and Briggs (1956) found that with increasing embryonic age, more and more transplanted nuclei displayed a loss of differentiation potential. This observation was confirmed by other investigators for several frog and salamander species (reviewed by McKinnell 1978, Di Berardino 1997a, 1997b). What made this observation particularly compelling was the fact that similar results were obtained by King and Briggs (1956) with R. Pipiens and by Gurdon (1974) with the South African clawed toad, X. Laevistwo very different anuran species. X. Laevis is primitive and aquatic, and it develops rapidly, whereas R. Pipiens is more evolutionary advanced and terrestrial much of the year, and it develops more slowly Yet despite the evolutionary differences between the animals, similar loss of nuclear potential was observed in the transplanted nuclei from both species (reviewed by McKinnell 1972).
Although endoderm cells of tailbud-stage R. Pipiens embryos have lost nuclear potential, Hennen (1970) was able to reverse these otherwise stable nuclear changes with technical modifications to the transplantation procedure. She added the polycationic amine, spermine, to complex chromatin proteins and lowered the environmental temperature to lengthen the cell cycle of the oocyte host. Of the complete blastulae that developed from enucleated egg cells into which tailbud-stage nuclei had been transplanted using these modifications, 62% developed into normal larvae. By contrast, when the modifications were not used, only 25% of the blastulae develops into normal larvae. Almost 30 years ago, Hennen (1970) concluded that if normal differentiation involves the selective repression of genetic information, then repression, however stable it might be under normal conditions, is reversible as far as nuclei from tailbud presumptive midgut are concerned. The implication of Hen-nen's work is that modifications in the nuclear transfer procedure can enhance the developmental expression of nuclei from advanced developmental stages.
Cloning of larval Ana adult nuclei:
Only a small minority of larval amphibian nuclei directed enucleated eggs to develop into fertile frogs: one gut nucleus from Pleurodeles prehatching larva (Aimar 1972) and, from Xenopus swimming larvae, 20 gut nuclei (Gurdon 1962), 2 intestinal nuclei (Gurdon and Uehlinger 1966), and 2 epidermal nuclei (Brun and Kobel 1972, Kobel et al. 1973). However, it is not known whether the few totipotent nuclei originated from differentiated cells or from contaminating stem cells. In contrast to the few totipotent amphibian nuclei from larval stages, no adult nuclei were found to be totipotent (reviewed by Di Berardino 1997a, 1997b).
Extensive nuclear transfer studies of differentiated larval and adult cells from R. Pipiens and X. Faevis revealed the nuclei of these cells to be multi-potent but not totipotent: Xenopus nuclei from melanophores, erythroblasts, skin, and lymphocytes injected into enucleated eggs directed the development of pre- or post-hatching tadpoles (reviewed by Di Berardino 1997a, 1997b). The most advanced tadpoles ensued from R. Pipiens erythrocyte nuclei from juvenile frogs: 7.8% directed the formation of feeding tadpoles that survived for up to a month (Di Berardino et al. 1986).
These studies are important for several reasons. One is that the terminally differentiated state was obvious by the shape and color of the donor red blood cell. The cell type of donor nuclei in a cloning experiment is not always known because of the complexity of biological tissues that are composed of both differentiated and stem cells. With mature erythrocytes, there can be no doubt as to the cell type because of their oval morphology and red color. Furthermore, the unique morphology of the mature amphibian nucleated blood cell is always associated with a particular phase of the cell cycle (G0, in this instance) and the virtual absence of transcriptional activity.
Another notable aspect of the study was that erythrocyte nuclei were first injected into metaphase I oocytes and conditioned for one day while the oocytes matured into metaphase II oocytes (the conventional host). The matured oocytes were then parthenogenetically activated by insertion of a glass needle, and the maternal (oocyte) nucleus was removed, leaving only the erythrocyte nucleus in the cytoplasm.
On the following day, blastulae had developed that then became nuclear donors for enucleated metaphase II oocytes. These cloned erythrocyte embryos developed into feeding larvae with hind limb buds (Figure 4). The use of metaphase I oocytes for conditioning the erythrocyte genome was based on previous studies, in which erythrocyte nuclei transferred to metaphase II oocytes failed to promote development of the host beyond the early gastrula stage; however, those nuclei exposed first to metaphase I and then metaphase II oocyte cytoplasm directed the hosts to develop into larvae (Di Berardino and Hoffner 1983).
These experiments were based on the hypothesis that molecular components of the oocyte cytoplasm that prepare oocyte chromosomes to participate in fertilization would similarly condition the genetic material of erythrocytes. Obviously, the developmental potential of these terminally differentiated erythrocyte nuclei was enhanced by this conditioning; however, the actual molecular mechanisms responsible for this nuclear reprogramming require investigation.
Finally, the experiments resulted in the most develop-mentally advanced cloned animals produced with adult nuclei before the advent of Dolly, the sheep cloned from an adult mammary gland cell by Wilmut et al. (1997). Like the larvae in the erythrocyte study, Dolly developed from a quiescent nucleus, one in the G0 part of the cell cycle. One cannot but ponder the significance of the quiescent phase associated with the successful results of these donor nuclei. Perhaps the successful results of these donor nuclei in the quiescent phase suggest that G0 nuclei integrate more normally into the cell cycle of the host than those in the other cell cycle phases (Campbell 1999).
Mammalian cloning:
Cloning of mammals via nuclear transfer was initially reported in mice during the early 1980s, approximately 30 years after the first tadpole clones were produced. Although there was considerable interest in extending cloning to mammalian species, these efforts were delayed until numerous technical parameters were modified for the small (approximately 100 µm in diameter) mammalian oocyte, including enucleation and in vitro culture of viviparous oocytes and embryos.
Investigators using mammalian cells were interested in the same fundamental question of nuclear potency during embryogenesis that was posed for cells of invertebrate and vertebrate species. In fact, a series of experiments involving blastomere separation and fusion, bisection of blastocysts, and injection of inner cell mass cells into the blastocyst had already demonstrated that totipotency is maintained in mammals until at least the blastocyst stage (reviewed in Di Berardino 1997a, 1997b). In addition to these basic studies, investigators seek to improve the genetic content of livestock species to benefit agriculture focused on nuclear transfer to oocytes of large domestic animals.
Nuclear transfers in mammals are performed primarily by cell fusion, using a procedure similar to the one originally developed for murine nuclear transplants (McGrath and Solter 1983) but with some modifications (Figure 5). A micropipette is inserted through the zona pellucida (i.e., the noncellular envelope surrounding the mammalian oocyte) and positioned over the oocyte's spindle; the micropipette is then used to withdraw the spindle, chromosomes, and first polar body.
Next, a micropipette containing an intact, genetically marked donor cell is inserted through the zona pellucida, and the cell is gently injected into the cavity between the oocyte's membrane and the zona pellucida. The two cells are usually fused by electro-fusion, whereby the electrical discharge causes breaks in the cell membranes of the oocyte and donor cell, permitting the contents of each to mingle before membrane healing. The electrical discharge can activate some oocytes, although in most cases, additional treatment is required.
Alternatively, in some mouse experiments, nuclei may be injected into oocytes that are subsequently activated (Wakayama et al. 1998, Wakayama and Yanagimachi 1999). The nuclear transplants are reared in vitro through various cleavage stages and then transferred to the uteri of surrogate mothers. Totipotency has been demonstrated for nuclei from various preimplantation stages of mouse and cattle (reviewed by Sun and Moor 1995, Di Berardino 1997a, 1997b, Fulka et al. 1998). In all cases, the clones developed into adults and produced normal progeny. So far, the only primates cloned by nuclear transfer are two rhesus monkeys (Figure 5) from 8-cell embryos (Meng et al. 1997). We await a report on tests of their fertility.
In some cases, multiple clones were produced from a single donor animal; these will be valuable for testing pharmaceuticals on animals with an identical genetic (nuclear) background. For example, isogenic groups were reported in murine twins and triplets (Kono et al. 1991) and in a group of over 30 animals produced by serial cloning (i.e., the cloning of clones; Park et al. 1993, Wakayama et al. 1998). Similarly, multiple calves ensued from one donor (Bondioli et al. 1990, Willadsen et al. 1991, Chesne et al. 1993), and 10 calves were produced by serial cloning (Stice and Keefer 1993). Finally, serial cloning of goat nuclei from the 32-cell stage resulted in 45 kids (Zang and Li 1998). Three identical transgenic goats were cloned from fetal cells bearing the human gene for antithrombin III, and one goat is producing the protein in her milk (Baguisi et al. 1999).
Although cloning of preimplantation mammalian nuclei has been relatively successful, the cloning efficiency from more advanced donor stages, as in amphibians, decreases. Nevertheless, a cloning efficiency adequate for commercial use has been achieved with fetal lamb and calf fibroblast and muscle cells, as well as with adult sheep mammary gland, mouse cumulus, and calf cumulus and oviductal cells.
The percentages of newborns, based on the number of embryo clones transferred to uteri, are 5-20% for lambs (7 total) from fetal fibroblast cells (Schnieke et al. 1997), 11% for calves (3 total) from fetal fibroblast cells (Cibelli et al. 1998), and 7% for calves (2 total) from fetal muscle cells (Vignon et al. 1998). With respect to the cloning efficiency from adult cells, the percentages are 3% for lambs (1 total) from mammary epithelium (Wilmut et al. 1997), 3% for mice (33 total) from cumulus cells (Wakayama et al. 1998), 1% for mice (3 total) from tail-tip cells (Wakayama and Yanagimachi 1999), 80% for calves (8 total) from cumulus and oviductal cells (Kato et al. 1998), and 10% for calves (10 total) from mural granulosa cells (Wells et al. 1999).
However, for those persons speculating on the application of cloning to humans, we emphasize the high mortality rate occurring throughout the experimental procedure. The Dolly experiment, as we will discuss subsequently, began with 434 attempts to fuse a mammary gland cell to an oocyte; the development of this one ewe represents a success rate of only 0.2%, and the remaining attempts resulted in death, either during fusion or during various pre- and postnatal stages. Although the success rates for mouse (1-2%) and calf (3.2%) cloning from adult cells are higher than for humans, the resulting mortality rate is still high.
Several studies in sheep and cattle deserve further discussion. Sims and First (1993) initially cloned four calves from cultured cells of the inner cell mass of blastocysts. This result established the feasibility of cloning from cultured cells, and it suggested that transgenic clones might be produced by transfecting cultured cells with a foreign gene and then using these cells as donor cells for nuclear transfer. Indeed, two laboratories have now produced transgenic cloned animals. Schnieke et al. (1997) cloned three transgenic lambs (two of which survived) containing the human gene for clotting factor IX. Fetal lamb cells in culture grew as fibroblasts and were transfected with constructs composed of the coding sequences for neomycin resistance and human clotting factor IX placed downstream of the promoter sequence for ovine ß-lactoglobin. The cultures were then exposed to G418, which kills all cells except those expressing neomycin resistance. Only those cells that had integrated the genes for neomycin resistance and, therefore, clotting factor IX survived and were used for nuclear transfer. The promoter sequence of the ovine ß-lactoglobin gene directs the gene for human clotting factor IX to be expressed in a tissue-specific manner, that is, in the mammary gland. After appropriate clinical trials, the protein harvested from the milk will be used to treat hemophiliacs (Wilmut 1998).
Recently, three transgenic calves were cloned from cells of a fetal fibroblastic cell line that contained a ß-galactosidase-neomycin resistance fusion gene driven by a constitutive promoter (Cibelli et al. 1998). In the future, transgenic clones can be designed to produce a variety of complex human proteins for human use. It should be noted that the production of transgenic animals by cloning is considerably more efficient than by gene injection into the pronuclei of fertilized eggs. There are several reasons for this increased efficiency, including the fact that transgenic cells can be selected in culture before nuclear transfer.
Previous Section Next Section:
Other biological problems examined by cloning:
Cloning experiments have provided valuable insight into a number of important cellular processes, such as nuclear reprogramming, cellular aging, and genomic imprinting.
Nuclear reprogramming:
The phrase nuclear reprogramming was used in frog cloning to designate the morphological and molecular changes occurring in nuclei transplanted into oocyte cytoplasm. The cytoplasm of activated eggs induces transplanted nuclei to cease RNA synthesis and synthesize DNA. The transplanted nuclei resume RNA synthesis at later embryonic stages, at the same time as embryonic nuclei from fertilized eggs begin to synthesize RNAs.
This reversibility of nuclear function also applies to specific genes (reviewed in Gurdon 1974). Moreover, during the first cell cycle of frog nuclear transplants, non-histone proteins move bidirectionally between the transplanted nucleus and the egg cytoplasm, whereas histone proteins primarily move from the cytoplasm into the nucleus (Di Berardino and Hoffner 1975, Hoffner and Di Berardino 1977). This result suggests that the chromatin proteins are being modified.
Today, techniques are available to analyze the remodeling of chromatin proteins directly. When transcriptionally inactive sperm nuclei were incubated in extracts from activated amphibian eggs, sperm-specific histone proteins were replaced by somatic histones H2A and H2B via the molecular chaperon nucleoplasmin (Katagiri and Ohsumi 1994). Similarly, erythrocyte chromatin was remodeled when the nuclei were incubated in extracts of activated amphibian eggs: somatic histones HI and Hl° were released from chromatin into the egg cytoplasm, oocyte-specific linker histone B4 and HMG1 were incorporated into remodeled chromatin, and somatic histones H2A and H4 were phosphorylated (Dimitrov and Wolffe 1996).
With respect to mammalian nuclei, recent studies in nuclear transplant embryos of mice, rabbit, pig, and cattle confirmed that changes similar to those observed in amphibian nuclear transplants also occur during nuclear reprogramming (Fulka et al. 1998). The importance of nuclear reprogramming is emphasized by the fact that incomplete nuclear reprogramming or its failure in amphibian and mammalian nuclear transplants causes abnormal and arrested development (reviewed in Di Berardino 1997a).
Scientists are only beginning to understand the molecular changes involved in nuclear reprogramming, yet this line of basic research may result in some of the most beneficial applications of cloning to humans. For example, if scientists could explain in molecular terms how a differentiated nucleus is de-differentiated, it might be possible to repair certain diseased tissuesa small amount of normal tissue could be removed from a patient and de-differentiated in culture.
After the cell population is expanded, appropriate inducers could be added to promote a desired type of cell differentiation (e.g., bone, cartilage, or muscle). Then, the tissue could be grafted to the patient's diseased areas, where the cells would be recognized as self and not rejected.
Cellular aging.
Cloning experiments have examined the replication potential of a genome during cellular aging. Normal cells cultured in vitro have a finite replication limit (Hayflick and Moorhead 1961). However, even after serial cloning through 145 cell cycles, nuclei of blastula cells were still able to direct tadpole development (Robert G. McKinnell, unpublished study). After serial transplantation to oocytes, even terminally differentiated erythrocyte nuclei that had gone through more than 110 cell cycles had the competence to direct the formation of tadpoles (Hoffner Orr et al. 1986). It is likely that somatic nuclei are rejuvenated to some large extent in oocytes because oocytes contain a large store of molecular substances that support nuclear replication and mitosis.
Further studies may reveal the mechanism of cellular rejuvenation. Telomeres, which are normally reduced in length during the aging process (Greider and Blackburn 1996, Shay 1997), may also be shortened in animal clones. Dolly and two other cloned sheep were reported to have telomeres that were shorter than those of age-matched controls (Shiels et al. 1999). Despite their shortened telomeres, the cloned sheep were vigorous and healthy. It remains to be seen if the reduced telomere lengths will have an effect during the lifetime of the sheep.
Genomic imprinting.
Nuclear transfer and molecular studies in mice elucidated genomic imprinting, a genetic mechanism that controls the differential expression of certain pairs of autosomal alleles. Mouse nuclear transplants constructed of two maternal or two paternal pronuclei fail to develop; only nuclear transplants comprising biparental nuclei form normal offspring (McGrath and Softer 1984, Surani et al. 1984). Molecular analyses of mouse embryos revealed that certain autosomal genes are differentially expressed from maternal and paternal genomes at specific times in development (Latham 1995).
Thus, an embryo with two inactive maternal (or paternal) genes fails to transcribe a necessary gene product. It is for this reason that the mammalian embryo requires a set of genes from both the father and the mother for normal development. The experimental results in mice led to the clarification of the basis of several inherited human disorders, such as Prader Willi and Angelman syndromes, which present different phenotypes but are both due to deletions in different homologues of chromosome 15: paternal deletions in Prader Willi patients and maternal deletions in Angelman patients (Driscoll 1994).
Previous SectionNext Section
Epilogue
Because of their fundamental nature, scientific discoveries in the basic sciences (e.g., anesthesia, atomic energy, recombinant DNA) occasionally lead to unanticipated deleterious applications. Knowledge in itself is amoral, but the choices for its applications reside in the ethical decisions of humans. Cloning, like other developments in basic science, was initiated to seek new fundamental knowledge. In addition to yielding information about the role of the nucleus during cell differentiation, the procedure also provided insight into basic aspects of modifying the cancer phenotype, rejuvenating aged nuclei by oocyte cytoplasm, nuclear reprogramming, and genomic imprinting. It will continue to yield new knowledge in these and other basic subjects.
As scientists who have worked in frog cloning, we are gratified to see decades of basic research culminating in the production of cloned mammals that will produce human proteins for the alleviation of human disease. Cloning will likely result in the genetic improvement of livestock, the production of animal models to study and treat human diseases, and sources of animal tissues and organs for xenotransplantation to humans. We consider these applications of cloning appropriate. Indeed, they are the reasons why Dolly was produced.
Once Dolly appeared, however, the news media became consumed with the idea of cloning human adults, which immediately stimulated a worldwide debate among ethicists, theologians, clerics, lawyers, legislators, and, of course, laypeople. We do not intend to summarize the many proposals and convictions of others; rather, we delineate briefly the reasons for our belief that human cloning is scientifically and ethically unsound. We define human cloning as the attempt to produce a human organism by any cloning procedure: blastomere isolation, bisection (splitting) of preimplantation embryos, and nuclear transplantation.
Nuclear transplantation of embryonic, fetal, or adult cells from all species results in abnormal animals at a frequency that increases with the age of the donor cell. The failures result from incomplete nuclear reprogramming and failed cell cycle matching, although other causes may in some cases be responsible. Abnormal nuclear transplants from all donor stages arrest development at various stages: nuclear transfer and activation, cleavage, organo-genesis, tadpole, and juvenile. In viviparous species, abnormal transplant embryos may fail to implant, and those that do implant may abort at various embryonic and fetal stages. Finally, those nuclear transplants that are born may die soon after birth or survive with birth defects.
Analyses of abnormal nuclear transplant embryos from frogs, mice, cattle, rabbits, and pigs revealed chromosomal and/or molecular abnormalities that could account for the morphological aberrations (reviewed in Di Berardino 1997a). Furthermore, even some normal nuclear transplant frog blastulae derived from embryonic or adult nuclei contained abnormal chromosomes in some of their cells, whereas the chromosomes in other cells of the same blastulae appeared normal. Such cases would pose confusion in monitoring normal-appearing nuclear transplants for further culture and development. For these reasons, we consider the use of any cell type (not just those from adults) for human cloning scientifically and ethically unsound.
In the case of Dolly, she was the only successful case out of 434 attempted fusions of oocytes and donor cells that were taken from cultures of mammary gland. Even as cloning of adult nuclei becomes much more efficient, there will still be hazards to humans. For example, the donor cells could suffer mutations in situ from radiation, chemicals, and/or aging during the lifetime of the donor. Mutations could also arise in the donor cells during cell culture, an event that is not unusual. There are still other scientific concerns.
Will telomere shortening in the donor cell limit the life span of the clone? Will stored gene products (RNAs and proteins) in oocytes from foreign donors always be compatible with the donor nucleus? Finally, it is important to consider that meiosis, which precedes sexual reproduction, affords humans another opportunity for DNA repair and therefore should not be avoided in favor of asexual (somatic) reproduction. For all of these reasons, we oppose human cloning.
Apollyon the Greek Sun god who was earlier worshipped during the Egyptian Eighteenth Dynasty of Akhenaten, the father of Tutankhamen, rises up on the earth in pursuit of global dominance and aggressively works to seize control over a final age of human apostasy. The Bastard child has grown up and serves in global prominence, waiting in the wings" for his reveal. Believers who pay attention to biblical prophecy know his identity and restrain him until they are taken out of the way... NOT by his hand but by the Hand of the Almighty One!
In 1907, the actual body of Akhenaten was discovered in Egypts Valley of the Kings by British archaeologist named Edward Ayrton. After unearthing Akhenatens mummified remains, Edward Ayrton was able to confirm that the ancient pharaohs skull was misshapen and elongated. Akhenaten was succeeded by his son, Tutankhamun, who became the most renowned pharaoh of all time. When his tomb was discovered in 1922 by Howard Carter, Tutankhamun was also found to have an elongated skull. In February 2010, the results of DNA tests confirmed that he was the son of Akhenaten. In 2012, , the tomb KV55 mummy was proven through DNA to be that of Akhenaten.
Amenhotep IV/Akhenaten - Birth name: Amenhotep ("Amun is satisfied"), Adopted name: (CHANGED HIS NAME) to Akhenaten/Akhenaton ("Servant of the Aten"), Throne name: Neferkheperure ("Beautiful are the Manifestations of Re"), Rule: 1350 - 1334 BC (10th king of the 18th dynasty, New Kingdom), Noteworthy relatives: Nefertiti (queen), Tutankhamun (son), Ankhesenamun (daughter), Merytaten (daughter), Smenkhkare (son), Amenhotep III (father), Tiy (mother).
He was born to Amenhotep III and his Chief Queen Tiyee and was their younger son. Akhenaten was not originally designated as the successor to the throne until the untimely death of his older brother, Thutmose. Amenhotep IV succeeded his father after Amenhotep III's death at the end of his 38-year reign, possibly after a short coregency lasting between either 1 to 2 years.
Pharaoh Akhenaten was known as the (RENEGADE), Heretic King. He was the tenth King of the 18th Dynasty. Egyptologists are still tying to figure out what actually happened during his lifetime as much of the truth was buried after he died.
Akhenaten lived at the peak of Egypt's imperial glory. Egypt had never been richer, more powerful, or more secure. Up and down the Nile, workers built hundreds of temples to pay homage to the Gods. They believed that if the Gods were pleased, Egypt would prosper. And so it did.
A religious reformer he made the Aten, the sun disc, the center of Egypts
Gem-pa-Aten temple at East Karnak - "the Aten is found" - Cairo Museum
O sole god without equal !
You are alone, shining in your form of the living Aten.
Risen, radiant, distant and near.
Great Hymn, 47 & 73-74.
SOURCES: Egyptologist Jan Assmann
Oxford Journals: http://bioscience.oxfordjournals.org/content/49/11/875.full
University of Maryland Medical Center: Akhenaten Deformities | University of Maryland Medical Center http://umm.edu/ /a-feminine-physique-a-long-thin-neck-and-e
http://umm.edu/ /a-feminine-physique-a-long-thin-neck-and-e
1 Professor of Genetics and Cell Biology at the University of Minnesota, Saint Paul, MN 55108-1095. He is the author of Cloning: Nuclear Transplantation in Amphibia (1978) and Cloning of Frogs, Mice and Other Animals (1985) and the 1998 recipient of the Prince Hitachi Prize in Comparative Oncology, awarded by the Japanese Foundation for Cancer Research. E-mail: mckinnel@biosci.umn.edu
2 Professor emerita of Biochemistry at MCP Hahnemann University, Philadelphia, PA 19129. She is the author of Genomic Potential of Differentiated Cells (1997) and the 1996 recipient of the Jean Brachet Memorial Award, given by the International Society of Differentiation, Inc..
http://beforeitsnews.com/ /youre-traveling-through-another-
http://www.livescience.com/39349-akhenaten.html
http://bioscience.oxfordjournals.org/content/49/11/875.full
http://conservativetribune.com/carson-criticizes-obama-se /
http://www.neferchichi.com/akhenaten.html
http://www.youtube.com/watch
http://www.heptune.com/Marfans.html
http://www.ancient-code.com/akhenaten-the-alien-pharaoh/
World leaders will announce that "Hyper Intelligent Alien" biological founders of the human race have provided indisputable evidence that proves that the origin of human development is the product of the ancestral inception of Alien male DNA. The Anti-Christ' father is the devil, he is called the "Son of Perdition", the offspring of "the father of ALL Lies"
Ancient Biblical history records the existence of "other beings" and their presence and sphere of influence throughout history on planet Earth. THE "BIG LIE" is that these entities are not "ALIEN" Hybrids from another planet but rather ANGELS, (fallen angels), who were expelled from the presence of God in ancient times. These creatures disguise themselves at "beings of light". Isaiah 14:12 / Luke 10:18 / Revelation 12:4.
Biblical History records that in ancient times, these "fallen beings" bred with some human women, creating a race of giants known as Nephilim. Nephilim embodied corrupt DNA and subsequently attempted at various intervals throughout human history, to physically dominate the Earth and its inhabitants. Both the fallen angels and their offspring the Nephilim, have wreaked havoc on earth every since the corruption of Adam. Read more:
Adam's corruption ran deep into his DNA. When Adam yielded himself to the Serpent, (Lucifer / Satan), who is the "commander and chief" of these "beings", the portal of another dimension, (Hell), literally opened onto the Earth and corruption entered into Adam and damaged the precious, Divinely designed and exclusive gift of incorruptible human DNA that held the unique genetic properties, "strands and markers", of life eternal.
Adam's broken relationship with his Creator ensured that he, Adam, would not live in his corrupted state of being forever. This is the very reason why a corrupt man needed a pure incorruptible God to intervene in this matter and provide a Divine Answer to satisfy the legal requirements rendering the eternal justice of a perfect God that would release man from the corruption of death.
Once the DNA was damaged, Satan gained access through man, to the entire Planet Earth, its world systems and future human race launching the "BIG LIE". Genesis 6:16
THE FINAL BIG LIE-
It will be announced in the near future that it was necessary to conceal the true identity of Barack Hussein Obama's "paternal" father in order to reveal to the world, at an appropriate moment, his true biological connection to ancient alien ancestry. 2 Cor 11:14. (It is the "illusion" of this claim that will make converts accept this final perverted global plot to rule the Earth.)
This announcement may occur sometime in the year 2025 and be followed by uncensored information from government sources revealing decades of "eighth kind" encounters and ongoing alien dialog with world leaders confirming this claim. It is at this time, with the assistance of Pope Francis that he will be received as a god sent to the inhabitants of Earth to save mankind.
In a final act of defiance against God, a last attempt to circumvent the Will of God and rule over planet Earth and inhabitants is now taking place in our generation.
The Truth is that God Himself became flesh, A virgin conceived the Seed of Jehovah and the Divine offspring, Messiah, Jesus The Christ was born to die in our place. The innocent for the guilty, the Lamb of God, slaughtered in the place of man, The Guilt Offering who took away the sins of all men and its sting of death for all who call upon His Name. John 3:16. HELL... THE (mAN of Lawlessness) may be the hideous creation of "ARTIFICIAL INSEMINATION" from the Ancient DNA of Akhenaten. THIS SON of PERDITION, "whose father IS the devil, carries out his father's desires. And like his father, who was a murderer from the beginning, not holding to the Truth, for there is no Truth in him. When he lies, he speaks his native language, for he is a liar like his father, the father of lies".
Hussein's fraudulent birth certificate was designed to not just cover "WHERE" he was born, but to ALSO not disclose "WHO" his genetic paternal, "father" honestly is. The "COVER-UP" will give credence to the future claim by Francis, the false prophet of Rome, that this "anti-Christ" is the "PROOF", (evidence) of the superior race of the extra-terrestrial genetic "fatherhood" an alien origin of human race.
The "man of lawlessness" is the appointed representative of an "extra-terrestrial" entity claim of human "origin" who has returned to earth to "save their own creation" from demise. As "man" will be presented to humanity, whose "paternal", (of the male genetic origin) is undocumented (not known) and whose mother, (maternal) genetic origin), is of a "whore" by nature, the exact opposite of Yeshua Messiah! This appointment by Hell will manifest at the time of the coming global government "alien among us disclosure". MOST of humanity will embrace the idea, thus Satan's GREAT DECEPTION, that will rob Heaven of scores of souls, who will follow this BEAST straight to Hell.
Terra firma (of earth) vs Extraterrestrial (Originating from outside of the Earth's atmosphere, from space, or from another planet; alien to Earth or its environment). Believers will need to cling to every Word of YHWH to overcome the lies that are manifesting about the human origin. Genesis specifically proclaims the "man" is the exclusive creation of YHWH taken from "TERRA FIRMA", (EARTH). Remember this when the proclamation is that man is from genetic alien extraterrestrial origin!!!
"the LORD God formed man out of the clay of the ground (terra firma) and blew into his nostrils the breath of life, and so man became a living being.
Akhenaten was an ancient Egyptian pharaoh and 10th ruler in the 18th dynasty. Professor Stuart Fleischmann and his team found some mutations in the genes of Akhenaten. The study resulted in a claim that Akhenaten's descendants are human-alien hybrids. The SAME claim will be made of the "son of perdition", (anti-Christ), once he takes his final global leadership role on behalf of satan. Obama's middle name was originally Muhammad rather than Hussein; that his mother had originally named him Barry rather than Barack; and that Barack Obama, Sr. was not his biological father, as well as the rumor that Barack Obama was not a natural-born Citizen. The official lie of Hussein Obamas birth that was presented by his handlers is that he was born on August 4, 1961, at the Kapiolani Maternity & Gynecological Hospital (now called the Kapiolani Medical Center for Women & Children) in Honolulu, Hawaii. His mother was Ann Dunham, from Wichita, Kansas, the true identity of his father is unknown. erronious information presented by the Dunham family regarding Hussein's birth is that an African, whose name is Barack Obama, Sr., a Luo from Nyangoma Kogelo in the Nyanza Province, in what was then the Colony and Protectorate of Kenya is his biological father. Sr. attended the University of Hawaii as a foreign student arranged by the Dunham family and was coerced into participating in a contrived "marriange" arrangement, under a threat of deportation by Stanley Armour Dunham, the father of Ann, a known CIA operative, in order to "cover Ann's bastardized pregancy by an unknown male.
Egyptian Pharaoh Akhenaten
"O'AKHENATEN ... OBAMA"
"O'AKHENATEN ... OBAMA"